| Research Article |
Open Access |
|
Imen Hammami1,2*, Afef Nahdi1, Souheila Amara2, Awatef Ben Ammar3,
Michèle V El May1 and Claire Mauduit2,4 |
| 1Research unity, Tunis 1007, Tunisia |
| 2University of Lyon, Oullins F-69921, France |
| 3Laboratory of Electronic Microscopy, Tunis 1007, Tunisia |
| 4Laboratory of Anatomy and Cytology-Pathology, Civil Hospitals of Lyon, Hospital Center of Lyon-Sud, Pierre-Benite cedex F-69495, France |
| *Corresponding author: |
Imen Hammami
Research unity no 01/UR/08-07
Faculty of Medicine
Tunis 1007, Tunisia
E-mail: hammamiimen@hotmail.fr |
|
| Â |
| Received April 20, 2012; Published October 29, 2012 |
| Â |
| Citation: Hammami I, Nahdi A, Amara S, Ammar AB, May MVE, et al. (2012) Germ Cells Apoptosis Induced by Chronic Garlic (Allium sativum) Feeding. 1:440. doi:10.4172/scientificreports.440 |
| Â |
| Copyright: © 2012 Hammami I, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. |
| Â |
| Abstract |
| Â |
| The aim of this study is to clarify the effects of crude garlic feeding (Allium sativum) on somatic and germ cells on adult rat. Twenty four adult male rats were divided into four groups: group 1 (untreated) and groups 2, 3 and 4 were fed for 30 days with 5 g, 10 g and 30 g/100 g of standard diet, respectively. Through TUNEL, immunostaining and Western blotting approaches, we showed that crude garlic feeding induced apoptosis in testicular germ cells (spermatocytes and spermatids) from rats fed with 10 g and 30 g of A. sativum. This cell death process was characterized by increased levels of active caspase-3 but not caspase-6. Expression of the caspase inhibitors c-IAP1 and c-IAP2 was increased at all doses of garlic, while expression of apoptosis inducing factor mitochondrionassociated (AIFM1) was unchanged. Moreover, expression of the IAP inhibitor Smac/DIABLO was increased from groups 3 and 4. In summary, we showed that feeding with crude garlic, for one month at high doses; induce apoptosis in testicular cells specially spermatocytes and spermatids but not somatic cells. |
| Â |
| Keywords |
| Â |
| Garlic; Germ cells; Apoptosis; Effector caspases; IAPs (Inhibitor of Apoptosis Proteins); AIF (Apoptosis Inducing Factor) |
| Â |
| Introduction |
| Â |
| Medicinal plants such as thyme, onion, blackseed, lemon, balm and nettle are intensively used in traditional medicine. Garlic or Allium sativum (A. sativum) is among the most important of these plants. Garlic is one of the ten commonest herbal medicines used in the United States (ranked in fifth place), according to recent sales data [1]. Indeed, A. sativum displays therapeutic effects such as in the treatment of hypercholesterolemia [2], prevention of arteriosclerosis [3] and some cancers [4], and presents anticoagulant [5] and antihypertensive [6] properties. For example, non-pharmacological treatment with garlic preparation is suggested to reduce blood pressure in hypertensive individuals [7]. The side effects, particularly on male reproduction, of such a chronic treatment are poorly investigated. To date, it has been reported that heated garlic juice was effective in recovery of testicular function after experimental testicular hypogonadism [8] but other laboratories have reported that powder [9] or crude [10] garlic preparations impaired testicular and male reproductive tract functions. Moreover, garlic metabolites such as diallyl trisulfide have been reported to have spermicidal effects [11,12]. The mechanisms of garlic action on male reproduction function, and particularly on spermatogenesis, however, remain unknown. In the present study, we tried to identify the cellular and molecular targets of crude garlic administrated in various doses to adult male rats. |
| Â |
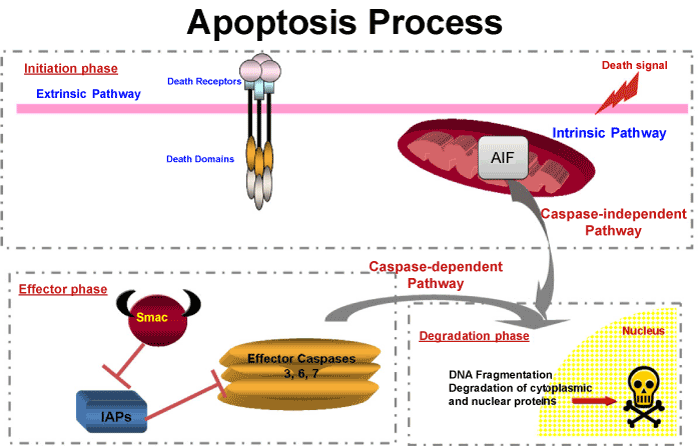
| In this work, we were interested to study the apoptosis on somatic and germ cells. We focused on the last step of apoptosis, before the irreversible cleavage of crucial proteins and endonuclease activation, which is under the control of effector caspases named Caspase 3, 6 or 7. Caspase 3 (CASP3) has been studied extensively and is known to be synthesized as a pro-enzyme which needs cleavage to be active [13]. Activation and/or activity of CASP3, 7 or 9 are regulated by the Inhibitors of Apoptosis Proteins (IAPs). IAPs share a common domain known as BIR (baculovirus IAP repeat), that binds to and inhibits CASP3, 7 or 9 [14,15] (Figure 1). Among the IAP family are XIAP, c-IAP1, c-IAP2 and Survivin [16]. A third level of regulation has also been observed. IAP action can be inhibited by mitochondrial proteins such as Smac/DIABLO [17-19]. Therefore, the fate of the cell at the executioner step of apoptosis depends on the relative quantity of each actor: effector caspase; IAPs and IAP inhibitors, such as Smac/DIABLO. In addition to the caspase pathway, the AIFM1 (Apoptosis inducing factor mitochondrion-associated 1) exists, which is a phylogenetically ancient mitochondrial intermembrane flavoprotein endowed with the unique capacity to induce caspase-independent peripheral chromatin condensation and large-scale DNA fragmentation when added to purified nuclei [20] (Figure 1). |
| Â |
|
|
Figure 1: Simplified diagram of apoptotic cell death process. |
|
| Â |
| Materials and Methods |
| Â |
| Plant and preparation |
| Â |
| The type of A. sativum used in the present study was “spring garlicâ€. This variety has pink bulbs and is planted between December and March (according to the weather) in Tunisia and collected in July. This type of garlic contains 2.1% proteins, 30% carbohydrates, 1.5% fibre, 0.2% fat, 0.015% vitamins and 0.7% minerals. The plant (A. sativum) used in this study was grown in Tunisia and purchased at a local market. Every day the garlic pellets were made by mixing peeled cloves of garlic with powdered standard rat pellet diet (Industrial society of food, Sfax, Tunisia) at four doses: 5 g, 10 g and 30 g/100 g of standard diet. For example, the last dose for one rat was prepared by mixing 9 g of crude garlic with 30 g of powdered standard diet in 5 mL of water. Cloves were crushed in distilled water to minimize volatile compound loss. A similar volume of water was added to the other doses. |
| Â |
| Animals and treatment |
| Â |
| A number of 24 adult male Wistar rats (Pasteur Institute of Tunis, Tunisia), whose average weight ranged between 200 g and 250 g, were used for the study. The animals were housed with proper aeration at 25 ± 2°C, and were given tap water ad libitum. The rats were allowed to acclimatize in the laboratory for a period of 1 week before the beginning of the study. |
| Â |
| The rats were randomly assigned into the different groups (of six animals each) using a hazard permutation table. Control animals received a standard pellet diet (group 1). The other groups received a diet supplemented with 5 g, 10 g and 30 g/100 g of standard diet (for groups 2, 3 and 4, respectively). All rats were weighed daily. After 30 days of treatment, rats were killed by decapitation. Testis were weighed and dissected. For each animal, the first testis was frozen at -80°C for Western Blotting analysis and the other was fixed in a 10% formaldehyde solution, for histo-pathological studies. |
| Â |
| All studies on animals were conducted in accordance with current regulations and standards approved by the Faculty of Medicine of Tunis animal care committee. |
| Â |
| TUNEL (TdT-mediated dUTP-X nick end labeling) |
| Â |
| Paraffin sections (5 μm) of formaldehyde-fixed testicular tissues were mounted onto Superfrost Plus slides. The sections were handled as previously described [21]. For each rat testis at least 100 random seminiferous tubules were numbered. The results were expressed as the apoptotic rate, i.e. the number of TUNEL positive cells per number of Sertoli cells. |
| Â |
| Immunohistochemistry |
| Â |
| Paraffin sections were incubated for 20 min at 93°–98°C in citric buffer (0.01 M, pH 6) and left to cool for 20 min at room temperature. The sections were rinsed twice for 5 min in osmosed water, and washed twice for 5 min in Tris buffered saline (TBS) containing 0.1% Tween-20. The Envision+ kit was used for detection of anti-cleaved CASP3 antibody (dilution 1/50) according to the manufacturer's recommendations. The antigen-antibody complexes were stained with DAB which generated a brown color at the site of peroxidase activity. The sections were rinsed twice for 5 min in osmosed water, counterstained with hematoxylin for 5 min and mounted in Faramount®. |
| Â |
| Western blotting analyses |
| Â |
| Proteins were obtained from testicular tissues as previously described [10]. Proteins (15-40 μg) were resolved on a 10%-15% sodium dodecyl sulfate/polyacrylamide gel. Proteins were electrophoretically transferred to a nitrocellulose membrane using 25 mM Tris-185 mM glycine buffer (pH 8.3) containing 20% methanol at a constant voltage of 100 V for 1 h. After transfer, the membranes were incubated in a blocking buffer (TBS containing 1% of BSA and 0.1% Tween-20) for 2 h at room temperature. The membranes were rinsed three times with TBS/0.1% Tween-20 for 10 min each, and incubated with the first antibody (in TBS containing 1% of BSA) overnight at 4°C. The antibodies were diluted as follows: 1/200 for CASP3; 1/400 for c-IAP1; 1/600 for Smac/DIABLO; 1/1,000 for CASP6 and 1/2,500 for c-IAP2. The protein loading was checked by probing the blot with a rabbit IgG anti-ACTIN antibody (1/20,000). The antigen-antibody complexes were detected with a chemiluminescent kit. The membranes were exposed on Biomax MR films. The intensity of the bands was determined with Opti-Quant software. The data were expressed as a target/actin protein ratio. |
| Â |
| Data analysis |
| Â |
| The results are expressed as the mean ± SD. For each condition, at least six different rats were used. A one-way analysis of variance (ANOVA) for independent groups was performed to determine whether there were differences between all groups and this was followed by the Bonferroni post hoc test at p |
| Â |
| Results |
| Â |
| TUNEL analysis in germ cells |
| Â |
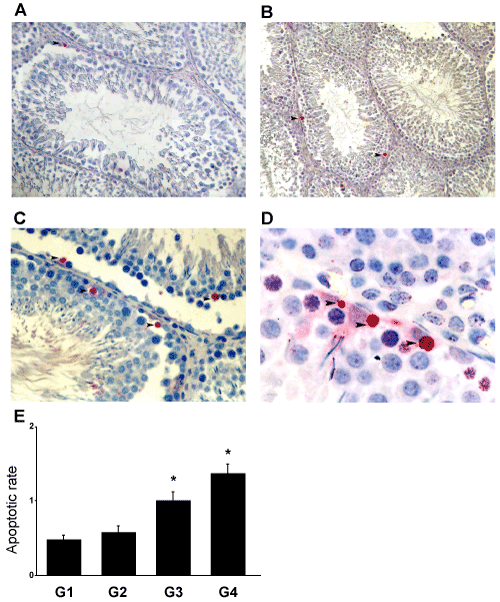
| Treatment with A. sativum induced a cell death process in the adult rat testis as shown by the TUNEL approach. In the control untreated animals, very few apoptotic germ cells were observed (Figure 2A), whereas TUNEL-positive cells were identified in rat testis treated with, 10 g (Figure 2B) and 30 g/100 g of standard diet (Figures 2C and 2D). These TUNEL-positive cells were mainly spermatocytes and spermatids (Figure 2D) but Sertoli and Leydig cells were not in touch with apoptosis. The number of apoptotic germ cells in rat testes increased after treatment with A. sativum in a dose-dependent manner (Figure 2E). A significant (p<0.05) increase was observed in the rats treated with 10 g and 30 g of A. sativum. |
| Â |
|
|
Figure 2: As induced apoptosis in germ cells. Detection of apoptotic cells was conducted through the TUNEL approach (A-D). (A) Testes from untreated rats (G1). (B) Testis of adult rat fed with garlic at doses 10 g (G3) (magnification= X40). (C and D) Testis of adult rat fed with 30 g of As (G4) (magnification= X40 and X100, respectively). (►) Red staining reveals the TUNEL positive nuclei in spermatocytes and spermatids in seminiferous tubules of adult rats. (E) The apoptotic rate represents the number of apoptotic germ cells (TUNEL approach) per Sertoli cells. The results are expressed as the mean ± SD. |
|
| Â |
| Detection of CASP3 in germ cells |
| |
| Â |
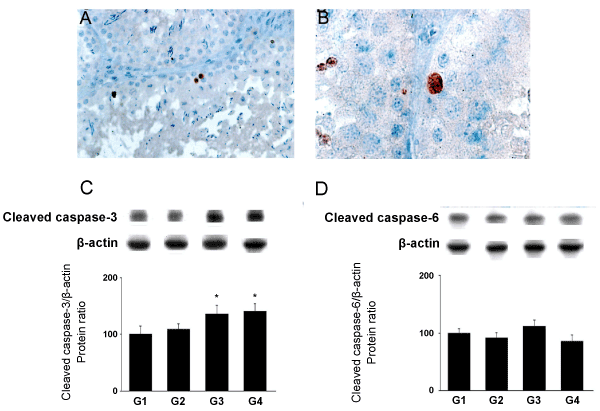
| By immunostaining, cleaved CASP3 was detected in spermatocytes and spermatids from rats fed with 10 g (G3; Figure 3A) and 30 g of A. sativum (G4; Figure 3B). There were no stained somatic cells detected in all rats. |
| Â |
|
|
Figure 3: Detection of cleaved CASP3 by immunostaining (A and B) and quantitative effects of As feeding on apoptosis in the testis by Western blotting (C and D).
Brown staining reveals the cleaved CASP3 cytoplasm immunostaining in testis of adult rat treated with garlic at doses 10g (G3) (A, magnification= X40) and 30g (G4) (B, magnification=X100). (C) Cleaved CASP3 and (D) cleaved CASP6 protein levels were analyzed through western blotting approach. The results are expressed as the mean ± SD. |
|
| Â |
| The cell death process induced in spermatocytes and spermatids from rats fed with A. sativum was probably an apoptotic mechanism, since cleaved CASP3 expression was increased in a dose dependent manner in the testicular tissues from rats fed with A. sativum with a significant increase in G3 (p<0.05) and G4 (p<0.05) (Figures 3C and 3D). In contrast, A. sativum -feeding did not modify the expression of cleaved CASP6 (Figure 2C). |
| Â |
| Effects of A. sativum feeding on IAPs expression in germ cells |
| Â |
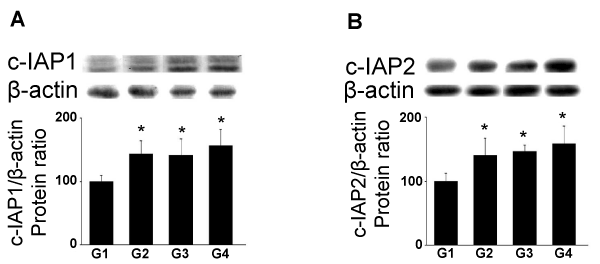
| The c-IAP1 protein levels were significantly increased in G2 (p<0.05), G4 (p<0.05) and G4 (p<0.01) of A. sativum (Figure 4A). Similarly, the c-IAP2 protein levels were significantly increased after treatment in G2 (p<0.05), G3 (31.5%; p<0.05) and G4 (37%; p<0.01) (Figure 4B). |
| Â |
|
|
Figure 4: Effects of As administration on c-IAP1 and c-IAP2 protein levels (Western blotting). Adult rats were untreated (G1) or fed with 5 (G2), 10 (G3) and 30g (G4) of As. (A) c-IAP1 and (B) c-IAP2 protein levels were analyzed through western blotting approach. In the upper panels, representative autoradiograms are shown. The results are expressed as the mean ± SD. |
|
| Â |
| Effects of garlic on mitochondrial pro-apoptotic factors expression |
| Â |
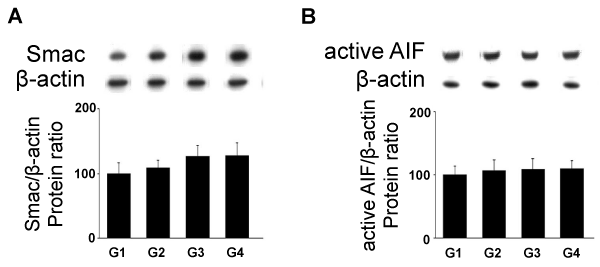
| The IAP inhibitor Smac/DIABLO protein levels were increased significantly in G3 (p<0.05) and G4 (p<0.05) (Figure 5A). The apoptotic cell death process can also be induced by a caspase-independent pathway represented by AIFM1. In the testicular tissue from rats fed with A. sativum, the expression of active AIFM1 protein (Figure 5B) levels was unchanged at the different tested doses. |
| Â |
|
|
Figure 5: Effects of As administration on mitochondrial pro-apoptotic factors expression (Western blotting). Adult rats were untreated (G1) or fed with 5 (G2), 10 (G3) and 30g (G4) of garlic. (A) Smac/DIABLO and (B) active AIFM1 protein levels were analyzed through western blotting approach. In the upper panels, representative autoradiograms are shown. The results are expressed as the mean ± SD. |
|
| Â |
| Discussion |
| Â |
| Garlic has acquired a reputation as a formidable prophylactic and therapeutic medicinal agent over the centuries and many favorable experimental and clinical effects of the consumption of garlic, in different types of preparations (crude, powder) have been reported (including cardiovascular diseases, stimulation of immune function, detoxification properties) [1]. It has long been known, however, that the extraction process can increase potency compared with the crude plant. In the present study, the chemical analysis of the crude garlic used was not achieved, but Shukla and Kalra [4] have quantified the concentration of the different compounds of garlic. In this context, administration of garlic preparation to prevent hypercholesterolemia or arteriosclerosis might have side-effects on other organs. In terms of testicular functions, garlic or its metabolites have been studied as a protective adjuvant to different types of toxins [22]. Indeed, induction of testicular hypogonadism by heat is prevented in part by different types of garlic preparation (raw, heated garlic juices, dehydrated garlic powder of the more potent aged garlic extract) [8]. Aqueous extract of garlic [23] or the metabolites diallyl sulfide [24] and diallyl tetrasulfide [25] offer a protection against cadmium-induced testicular damages. Garlic is also effective in restoring the testicular histology altered by EDTA [26]. The antioxidant activities of garlic extract were shown to decrease the toxic effects of free radicals induced by testicular torsion and detorsion [27]. While several studies show that A. sativum has a protective effect as an adjuvant, other studies show that it modifies spermatogenesis. Indeed, daily administration of 50 mg garlic powder over 70 days induced a spermatogenetic arrest at spermatocyte I stage [9]. Moreover, aqueous garlic extract [11] or the metabolite diallyl trisulfide [12] has spermicidal effects. In contrast, a 90-day administration of 100 mg/kg aqueous extract of garlic has been shown to increase the number of spermatozoa [28]. In this context, to assess the positive or negative effect of garlic, it was important to analyze its action on the different testicular cells through the study of germ and somatic cell apoptosis. |
| Â |
| In this context, this study has focused on the effects of A. sativum on testicular cells. We showed here that chronic crude garlic feeding induced germ cell death via an apoptotic process. This cell germ cell apoptotic process target spermatocytes and spermatids, whereas spermatogonia and somatic Leydig and Sertoli cells were not affected. Given that CASP3 was immunodetected mainly in spermatocyte and spermatid cells (the TUNEL-positive germ cells) and that garlic treatment, at the same doses as observed for TUNEL, induced a dose-dependent increase in active CASP3 protein levels, the possibility exists that the germ cell apoptotic process could be related to CASP3 activation. The germ cell death might also, however, be related to another programmed cell death process since CASP3 activation has been associated with differentiation in some cell types [29]. In contrast, the active effector CASP6 seemed not to be involved in our model, while proCASP6 is expressed in normal germ cells [30]. While it was previously reported that chronic administration of 50 mg of garlic powder induced an arrest of spermatogenesis, our present study shows, for the first time, the involvement of an apoptotic process that targets testicular germ cells. Very few, if any, studies have reported such an apoptotic effect of A. sativum on non-tumoral cells, although it has been described in tumoral cells [31]. For example, administration of crude extract of garlic to a human colon cancer cell line induced apoptosis by increasing the levels of BAX, CYCS (previously known as Cytochrome c) and CASP3 activity while it decreased the mitochondrial membrane potential [32]. More specifically, raw crushed garlic is high in allicin, a powerful bioactive compound of garlic that induced activation of CASP3, CASP8 and CASP9 and cleavage of poly (ADP-ribose) polymerase in several cancer cells [33]. Given that the balance between pro-apoptotic molecules (active effector caspase, IAPs inhibitors) and anti-apoptotic molecules (IAPs) levels determines the fate of the cells towards the executioner step of the death process [15], we have evaluated the expression of IAPs in our experimental model. C-IAP-1 and c-IAP2 are highly expressed in rat testes [34]. C-IAP-1 was detected in the nucleus of spermatogonia B, spermatocytes at different stages and of somatic cells, while c-IAP2 was localized in the cytoplasmic compartment of spermatocytes (from stage VI pachytene onwards), spermatids (round and elongated) and Leydig cells [35]. While c-IAP-1 and c-IAP2 play a role in the protection of germ cells from Fas-mediated apoptosis [34], their increased expression after treatment with crude garlic (the present study) suggested that germ cells are unable to inhibit CASP3 action. A potential explanation is that the high levels of Smac/DIABLO (expressed in spermatocytes) or HTRA2 in spermatids by inhibiting the IAP action may favor the activity of CASP3 in the germ cells mainly affected by apoptosis after A. sativum treatment. |
| Â |
| Crude garlic consumption by humans ranges from one to two cloves (about 4 g) to 28 g per day [1]. The doses used in the present study exceeds this amount of consumption, but various other types of garlic preparation are consumed (such as garlic powder, oil, extract or aged garlic) and the concentration in garlic active components is highly variable, particularly in powder and oil [1]. Moreover, the bioactive components of garlic are not fully characterized even if it is assumed that the sulfur-containing molecules are the active ones. Another point is that garlic consumption to reduce cardiovascular risk is a long-term consumption which is another potential negative effect with regard to spermatogenesis. In this context, and more widely, extrapolation from rat to human is a difficult task. It has long been known, however, that human spermatogenesis is more sensitive to stress than that of rats [36], suggesting that concentrations lower than those used in the present study might impair male spermatogenesis and, particularly, might induce azoospermia in men with low sperm count. |
| Â |
| Conclusion |
| Â |
| In summary, we showed that feeding with crude garlic, at high doses, for one month has inhibitory effects on spermatocytes and spermatids cells via an apoptotic process but Sertoli and Leydig cells were not affected by this process. |
| Â |
| |
| References |
| Â |
- Amagase H, Petesch BL, Matsuura H, Kasuga S, Itakura Y (2001) Intake of garlic and its bioactive components. J Nutr131: 955S-962S.
- Mahmoodi M, Islami MR, Asadi Karam GR, Khaksari M, Sahebghadam Lotfi A, et al. (2006) Study of the effects of raw garlic consumption on the level of lipids and other blood biochemical factors in hyperlipidemic individuals. Pak J Pharm Sci 19: 295-298.
- Siegel G, Klussendorf D (2000) The anti-atheroslerotic effect of Allium sativum: statistics re-evaluated. Atherosclerosis 150: 437-438.
- Shukla Y, Kalra N (2007) Cancer chemoprevention with garlic and its constituents. Cancer Lett247: 167-181.
- Steiner M, Li W (2001) Aged garlic extract, a modulator of cardiovascular risk factors: a dose-finding study on the effects of AGE on platelet functions. J Nutr 131: 980S-984S.
- Silagy CA, Neil HA (1994) A meta-analysis of the effect of garlic on blood pressure. J Hypertens 12: 463-468.
- Ried K, Frank OR, Stocks NP, Fakler P, Sullivan T (2008) Effect of garlic on blood pressure: a systematic review and meta-analysis. BMC Cardiovasc Disord8: 13.
- Kasuga S, Uda N, Kyo E, Ushijima M, Morihara N, Itakura Y (2001) Pharmacologic activities of aged garlic extract in comparison with other garlic preparations. J Nutr131: 1080S-1084S.
- Dixit VP, Joshi S (1982) Effects of chronic administration of garlic (Allium sativum Linn) on testicular function. Indian J Exp Biol 20: 534-536.
- Hammami I, Nahdi A, Mauduit C, Benahmed M, Amri M, et al. (2008) The inhibitory effects on adult male reproductive functions of crude garlic (Allium sativum) feeding. Asian J Androl 10: 593-601.
- Chakrabarti K, Pal S, Bhattacharyya AK (2003) Sperm immobilization activity of Allium sativum L. and other plant extracts. Asian J Androl5: 131-135.
- Qian YX, Shen PJ, Xu RY, Liu GM, Yang HQ, et al. (1986) Spermicidal effect in vitro by the active principle of garlic. Contraception34: 295-302.
- Hengartner MO (2000) The biochemistry of apoptosis. Nature407: 770-776.
- Clem RJ, Miller LK (1994) Control of programmed cell death by the baculovirus genes p35 and iap. Mol Cell Biol14: 5212-5222.
- Liston P, Fong WG, Korneluk RG (2003) The inhibitors of apoptosis: there is more to life than Bcl2. Oncogene 22: 8568-8580.
- Vaux DL, Silke J (2005) IAPs, RINGs and ubiquitylation. Nat Rev Mol Cell Biol6: 287-297.
- Liston P, Fong WG, Kelly NL, Toji S, Miyazaki T, et al. (2001) Identification of XAF1 as an antagonist of XIAP anti-Caspase activity. Nat Cell Biol 3: 128-133.
- Liu Z, Sun C, Olejniczak ET, Meadows RP, Betz SF, et al. (2000) Structural basis for binding of Smac/DIABLO to the XIAP BIR3 domain. Nature 408: 1004-1008.
- Wu G, Chai J, Suber TL, Wu JW, Du C, et al. (2000) Structural basis of IAP recognition by Smac/DIABLO. Nature408: 1008-1012.
- Cande C, Cecconi F, Dessen P, Kroemer G (2002) Apoptosis-inducing factor (AIF): key to the conserved caspase-independent pathways of cell death? J Cell Sci115: 4727-4734.
- Mauduit C, Siah A, Foch M, Chapet O, Clippe S, et al. (2001) Differential expression of growth factors in irradiated mouse testes. Int J Radiat Oncol Biol Phys50: 203-212.
- Oi Y, Imafuku M, Shishido C, Kominato Y, Nishimura S, et al. (2001) Garlic supplementation increases testicular testosterone and decreases plasma corticosterone in rats fed a high protein diet. J Nutr 131: 2150-2156.
- Ola-Mudathir KF, Suru SM, Fafunso MA, Obioha UE, Faremi TY (2008) Protective roles of onion and garlic extracts on cadmium-induced changes in sperm characteristics and testicular oxidative damage in rats. Food Chem Toxicol46: 3604-3611.
- Sadik NAH (2008) Effects of diallyl sulfide and zinc on testicular steroidogenesis in cadmium-treated male rats. J Biochem Mol Toxicol22: 345-353.
- Murugavel P, Pari L (2011) Diallyl tetrasulfide modulates the cadmium- induced impairment of membrane bound enzymes in rats. J Basic Clin Physiol Pharmacol18: 37-48.
- Khalil WK, Ahmed KA, Park MH, Kim YT, Park HH, Abdel-Wahhab MA (2008) The inhibitory effects of garlic and Panax ginseng extract standardized with ginsenoside Rg3 on the genotoxicity, biochemical, and histological changes induced by ethylenediaminetetraacetic acid in male rats. Arch Toxicol 82: 183-195.
- Unsal A, Eroglu M, Avci A, Cimentepe E, Guven C, et al. (2006) Protective role of natural antioxidant supplementation on testicular tissue after testicular torsion and detorsion. Scand J Urol Nephrol40: 17-22.
- al-Bekairi AM, Shah AH, Qureshi S (1990) Effect of Allium sativum on epididymal spermatozoa, estradiol-treated mice and general toxicity. J Ethnopharmacol29: 117-125.
- Miura M, Chen XD, Allen MR, Bi Y, Gronthos S, et al. (2004) A crucial role of caspase-3 in osteogenic differentiation of bone marrow stromal stem cells. J Clin Invest114: 1704-1713.
- Omezzine A, Chater S, Mauduit C, Florin A, Tabone E, et al. (2003) Long-term apoptotic cell death process with increased expression and activation of caspase-3 and -6 in adult rat germ cells exposed in utero to flutamide. Endocrinology 144: 648-661.
- Xiao D, Choi S, Johnson DE, Vogel VG, Johnson CS, et al. (2004) Diallyl trisulfide-induced apoptosis in human prostate cancer cells involves c-Jun N-terminal kinase and extracellular-signal regulated kinase-mediated phosphorylation of Bcl-2. Oncogene 23: 5594-5606.
- Su CC, Chen GW, Tan TW, Lin JG, Chung JG (2006) Crude extract of garlic induced caspase-3 gene expression leading to apoptosis in human colon cancer cells. In Vivo20: 85-90.
- Oommen S, Anto RJ, Srinivas G, Karunagaran D (2004) Allicin (from garlic) induces caspase-mediated apoptosis in cancer cells. Eur J Pharmacol 485: 97-103.
- Holcik M, Lefebvre CA, Hicks K, Korneluk RG (2002) Cloning and characterization of the rat homologues of the Inhibitor of Apoptosis protein 1, 2, and 3 genes. BMC Genomics3: 5.
- Wang Y, Suominen JS, Parvinen M, Rivero-Muller A, Kiiveri S, et al. (2005) The regulated expression of c-IAP1 and c-IAP2 during the rat seminiferous epithelial cycle plays a role in the protection of germ cells from Fas-mediated apoptosis. Mol Cell Endocrinol245: 111-120.
- Meistrich ML, Samuels RC (1985) Reduction in sperm levels after testicular irradiation of the mouse: a comparison with man. Radiat Res102: 138-147.
|
| Â |
| Â |