| Research Article |
Open Access |
|
| G Mabilleau, R Gill and A Sabokbar* |
| Institute of Musculoskeletal Sciences – The Botnar Research Centre, Nuffield Department of Orthopaedics, Rheumatology and Musculoskeletal Science, University of Oxford, OX3 7LD, UK |
| *Corresponding authors: |
Dr Afsie Sabokbar
The Botnar Research Centre
Windmill road, Oxford OX3 7LD, UK
E-mail: afsie.sabokbar@ndorms.ox.ac.uk |
|
| |
| Received February 17, 2012; Published June 27, 2012 |
| |
| Citation: Mabilleau G, Gill R, Sabokbar A (2012) Cobalt and Chromium Ions Affect Human Osteoclast and Human Osteoblast Physiology In vitro. 1: 219. doi:10.4172/scientificreports.219 |
| |
| Copyright: © 2012 Mabilleau G, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. |
| |
| Abstract |
| |
| Introduction: Cobalt-Chromium (Co-Cr) alloys are widely used in biomedicine owing to their resistance to corrosion, mechanical properties, and biocompatibility. Despite the increase in its use in orthopaedic surgery, a number of unique complications (elevated serum levels of Co and Cr ions, development of pseudotumour, neck narrowing, osteolysis and fracture) have been recently reported with Metal-on-Metal (MoM) bearings. The goal of the present study was to assess the effects of Co and Cr ions on human osteoblasts and human osteoclast biology. |
| ` |
| Materials and methods: Peripheral blood mononuclear cells (PBMCs) have been used as a source of osteoclast precursors whilst SaOS-2 cells have been used as osteoblasts. Both cell types have been cultured in the presence of Co2+ or Cr3+ ions. |
| |
| Results: We have found that 100 μM Co2++ induced a significant increase in the number and the size of multinucleated tartrate resistant acid phosphatase positive osteoclasts as well as the number of nuclei per osteoclast. In contrast, 100 μM Cr3+ significantly decreased the number of newly-formed osteoclast but had no effect on the size or the number of nuclei per osteoclast. Interestingly, 100 μM of Co2+ was capable of strongly inhibiting bone resorption whereas 100 μM of Cr3+ had no significant effects on this parameter. The alkaline phosphatase activity was significantly increased by low concentrations of Co2+ and decreased by high concentrations of Cr3+ after 24 h and 48 h. Moreover, the degree of mineralization of a new bone matrix in vitro was significantly reduced when the SaOS-2 cells were exposed to high concentrations of Cr3+, but significantly increased when they were exposed to Co2+. |
| |
| Discussion: These results suggest that long-term exposure to “chronic” levels of metal ions damages the functional behaviour of bone cells and subsequently may affect bone quality. |
| |
| Keywords |
| |
| Osteoclast; Osteoblast; Cobalt ion; Chromium ion; Resurfacing; Metal-on-metal |
| |
| Introduction |
| |
| Cobalt-chrome (Co-Cr) alloys are widely used in biomedicine owing to their resistance to corrosion, mechanical properties, and biocompatibility. Co-Cr alloys are preferred in orthopaedics to nitinol and titanium alloys due to improved strength, wear resistance, toxicity and cost. Recently, in order to reduce the amount of wear debris produced by joint replacements, the use of new surface bearings, such as metal-on-metal (MoM), made of Co-Cr alloys, has rapidly increased. MoM hip resurfacing has been an attractive option for treating young and active patients with hip osteoarthritis. The perceived advantages of MoM bearings, mainly manufactured in Co-Cr alloys, include low wear rate, bone conservation, increased mobility and low dislocation rate (due to the large head size of the implant). |
| |
| Despite an increase in the use of MoM bearing implants in orthopaedic surgery, some concerns are raised especially regarding the serum levels of metal ions. Recent studies have reported that patients, undergoing MoM resurfacing, have elevated serum levels of Co and Cr ions [1,2]. The synovial fluids levels of these metal ions have been reported to be markedly higher in MoM patients where measured levels of Co and Cr were increased by 300-fold and 1000-fold respectively compared with serum levels [3,4] with unknown consequences for bone properties. Nowadays, it is widely accepted that bone strength depends on both bone quantity and “quality”, a term encompassing structural and material properties. Recently, we reported that Co and Cr ions affected the physical properties of hydroxyapatite crystals and could contribute to decrease the biomechanical strength of the mineral [5]. During physiological conditions, bone resorption and bone formation are closely related. Although osteolysis has also been associated with the early failure of MoM joint replacement [6-8], it is a rare phenomenon which occurs at a lower rate than that reported for patients with polyethylene implants. However, it is important to understand what are the effects of high metal ion concentrations on osteoclasts, the bone-resorbing cells and osteoblasts, the bone-forming cells. Despite previous studies investigating the effects of metal ions on rodent osteoclasts [9,10], little is known about the capacity of human circulating osteoclast precursors to transform into bone-resorbing osteoclasts in the presence of Co and Cr ions. Similarly, few studies have investigated the capacity of metal ions to induce osteoblastic cell death or alteration of osteoblastic metabolism [11,12]. However, these studies have been conducted in rodent cells or with concentrations higher than those commonly found in joint fluids of patients with MoM implants. As such, it is poorly understood whether in the presence of metal ions, osteoblasts can produce and mineralize a new collagen matrix. |
| |
| The aims of the present study was to evaluate the direct effects of Co and Cr ions on osteoclast precursors isolated from healthy volunteers and osteoblasts by assessing the cell viability, extent of osteoclast formation and bone resorption and synthesis and mineralisation of a new matrix in vitro. |
| |
| Materials and Methods |
| |
| Metal ions preparation |
| |
| To evaluate the effects of metal ions in vitro, CoCl2 and CrCl3 were purchased from Sigma – Aldrich Chemicals (Poole, UK) and dissolved in sterile phosphate buffer saline. The range of Co2+ and Cr3+ concentrations used to assess the extent of osteoclastogenesis, viability and cytokine release in vitro, were 1, 10 and 100 μM unless otherwise specified. These concentrations were chosen based on the recent findings reported by De Smet et al. [3]. The concentrations of Cl- in PBS were adjusted in order to avoid a dramatic increase in this element after dissolution of the metal salt. |
| |
| Cell culture |
| |
| To evaluate the response of normal osteoclast precursors to Co and Cr ions, peripheral blood mononuclear cells (PBMCs) were isolated as described previously [13] from the blood of 8 healthy volunteers (National Blood Service, Bristol, UK) with no history of metal allergy. Briefly, blood was diluted 1:1 in α-minimal essential medium (MEM) (Invitrogen, Paisley, UK), layered over Histopaque (Sigma-Aldrich Chemicals, Poole, UK), and centrifuged (693 x g) for 20 minutes. The interface layer was removed and resuspended in MEM supplemented with 10% heat-inactivated foetal calf serum (FCS, Invitrogen, Paisley, UK). A fraction of the cell suspension was counted in a haemocytometer following lysis of red blood cells by a 5% (v/v) acetic acid solution. |
| |
| To evaluate the response of human osteoblasts to metal ions, osteoblast-like cells SaOS-2 (American Type Culture Collection ATCC, Teddington, UK) were cultured in MEM supplemented with 10% heat- inactivated foetal calf serum, 100 UI/ml penicillin and 100 μg/ml streptomycin in a humidified atmosphere of 5% CO2 at 37°C. |
| |
| Viability in response to Co2+and Cr3+ |
| |
| As long-term cultures of human PBMCs in the absence of survival factor (i.e. Macrophage-Colony Stimulating Factor) is not feasible in vitro, the effect of viability and release of inflammatory cytokines in response to Co2+ and Cr3+ were only assessed in 24 h and 48 h of cultures. For this purpose, 1x105 PBMCs from healthy volunteers and 1x105 SaOS-2 cells were cultured in the presence or absence of Co2+ and Cr3+. The cell number was assessed using the MTT assay [3-(4,5 Dimethyl-thiazol-2-yl) 2,5-diphenyl tetrazolium bromide] as described previously [14]. |
| |
| Extent of osteoclast formation and bone resorption in response to metal ions |
| |
| To assess the extent of osteoclast formation and activation in response to metal ions, human PBMCs were cultured on glass coverslips and dentine slices as described previously [15] with or without Co2+or Cr3+. All cultures were maintained in the presence of 25 ng/ml recombinant human macrophage colony stimulating factor (M-CSF; R&D Systems Europe, Abingdon, UK), and 100 ng/ml recombinant human soluble receptor activator of nuclear factor kB ligand (sRANKL; Pepro-Tech EC Ltd. London, UK), for up to 21 days. |
| |
| After 14 days in culture, the coverslips were examined histochemically for the expression of tartrate resistant acid phosphatase (TRACP), one of the osteoclastic markers, as previously described [16]. Coverslips were then counterstained with 4’,6’-diaminido-2- phenylindole for 20 minutes and TRACP positive cells, with more than three nuclei, were identified as osteoclasts. The number of newly generated osteoclasts and the number of nuclei per osteoclast were assessed using light and fluorescent microscopic examination. The size of the osteoclasts and intracellular TRACP level were determined by image analysis using the Image J Freeware (NIH, Bethesda, MD, https:// rsbweb.nih.gov/ij/). |
| |
| After 18 days, the expression of osteoclast-associated receptor (OSCAR) and calcitonin receptor (CT-R) was assessed by Western blot as described previously [15] using specific antibodies for OSCAR (clone D19) and CT-R (clone N20) (Santa Cruz Biotechnology, Heidelberg, Germany). |
| |
| After 21 days in culture, the dentine slices were placed in NH4OH (1N) for 30 minutes and sonicated for 5 minutes to remove any adherent cells. After rinsing in distilled water, the dentine slices were stained with 0.5% (v/v) toluidine blue prior to examination by light microscopy. The surface of each dentine slice was examined for evidence of lacunar resorption. The extent of eroded surface was determined by image analysis on each dentine slice and expressed as the percentage of surface area resorbed per osteoclast. |
| |
| Oxidative stress measurement |
| |
| In order to determine the effects of metal ions on the extent of oxidative stress in osteoblasts in vitro, SaOS-2 cells were plated at a density of 2x104 cells/cm2 and cultured for 8 hours in the presence of 1, 10 and 100 μM metal ions. Intracellular levels of reactive oxygen species were determined using dichlorofluorescein diacetate (DCFH-DA) as previously described [16]. |
| |
| Alkaline phosphatase assay |
| |
| SaOS-2 cells were plated at a density of 2x104 cells/cm2 and cultured for 24 h and 48 h in the presence of 100 μM metal ions. Cells were washed in phosphate buffer saline and freeze-thawed 3 times, and alkaline phosphatase (ALP) was released by scraping the cells with 0.2% Nonidet P-40. ALP activity (pmol/ml of material) was determined using the fluorescent substrate 4-methyl umbelliferyl phosphate as previously described [17]. The yield of ALP was calculated by the preparation of a standard curve of 0-1000 pmol of 4-methylumbelliferyl phosphate on the same plate and standardised against the total amount of protein/ well, determined using a BCA protein assay kit (Pierce, Cramlington, UK). |
| |
| Mineralisation assay |
| |
| SaOS-2 cells were cultured for 21 days in the presence of 50 μg/ ml ascorbic acid (AA), 10 mM β−glycerophosphate (GP) and 100 μM metal ions. The formation of calcium phosphate in the culture was determined using alizarin red-S assay. Briefly, cells were washed with PBS, fixed in 100% ethanol (40°C, 1 hour), washed in water and stained with alizarin red-S (40 mM solution, pH 4.2) at room temperature under moderate shaking for 10 minutes. Cells were then washed with PBS. Photographs of each well were taken prior to incubation with 100 μL/ cm2 of 10% (w/v) cetylpyridinium chloride, 10 mM sodium phosphate, pH 7.0, at room temperature for 15 minutes under moderate shaking. Aliquots of the resulting supernatants were diluted 10-fold with the use of this buffer and the absorbance was measured at 560 nm. Values were normalized to total number of cells determined in parallel cultures. |
| |
| Statistical analysis |
| |
| All in vitro experiments were repeated at least 6 times. Results were expressed as mean ± standard error of the mean. The non parametric Kruskall-Wallis test was used to compare the differences between the groups. When significant differences were observed with respect to ion concentration, data were subjected to Mann-Whitney U test. Differences at p < 0.05 were considered significant. |
| |
| Results |
| |
| Co2+ and Cr3+ do not affect the cell number |
| |
| Adherent human PBMCs exposed to Co2+ or Cr3+ showed no significant differences in viability after 24 h and 48 h compared to control cultures (Figure 1A). SaOS-2 cells exposed to Co2+ for 24 h and 48 h showed no significant differences in viability compared to untreated cultures (Figure 1B). On the other hand, SaOS-2 exposed to Cr3+ exhibited a dose-dependent decrease of the cell number after 24 h and 48 h compared to control cultures but these differences did not reach statistical significance. |
| |
|
|
Figure 1: Assessment of human circulating osteoclast precursor number (A) and human osteoblast number (B) after 24h and 48h exposure to Co2+ and Cr3+. White bar = untreated cultures, black bars=Co2+- treated cultures and grey bars=Cr3+-treated cultures. |
|
| |
| Cytokin release |
| |
| |
| As shown in Figure 2A, PBMCs exposed to 100 μM Co2+, significantly release more IL-1β, IL-6 and VEGF as compared with untreated cells. On the other hand, PBMCs exposed to 100 μM Cr3+, significantly increased the release of TNF-α, IL-1β and MIP-1α (Figure 2B). |
| |
|
|
Figure 2: Cytokine release by PBMCs in response to 100μM Co2+ (A) and Cr3+ (B) after 24h and 48h cultures. Dash line represents the level of pro-inflammatory cytokine release in untreated cells. *: p<0.05 compared to untreated PBMCs. |
|
| |
| Extent of osteoclast formation |
| |
| Figure 2A shows an example of newly-formed multinucleated TRACP+ giant cells obtained in vitro in the presence of M-CSF and RANKL ± Co2+. In the absence of sRANKL, no multinucleated TRACP positive cells were noted on coverslips (data not shown). The multinucleated TRAP positive cells obtained with M-CSF and RANKL expressed OSCAR and CT-R, two markers expressed only by differentiated osteoclasts. Multinucleated cells generated in the presence of Co2+ and Cr3+ expressed also OSCAR and CT-R indicating that these multinucleated giant cells are also osteoclasts. Compared to control cultures (i.e. M-CSF + sRANKL only), 1 μM and 10 μM of Co2+ and Cr3+ had no effect on the number of the newly-generated osteoclasts (Figure 2B). However, at higher concentrations, the response differed based on the nature of the metal ions; compared to control cultures, 100 μM of Co2+ significantly increased the differentiation of osteoclast precursors into multinucleated osteoclasts as evidenced by a 5- fold increase in the absolute number of osteoclasts formed in vitro (174 ± 3 vs. 35 ± 5; p<0.0001). However, 100 μM of Cr3+ clearly hampered the osteoclastic differentiation, as a significant reduction in the absolute number of newly formed osteoclasts was evident (13 ± 4 vs. 35 ± 5 p<0.0001) (Figure 2B). |
| |
| Characterisation of newly-formed osteoclasts |
| |
| The number of nuclei per osteoclast, which reflects how many osteoclast precursors fused together to form multinucleated osteoclasts, was significantly increased only at the high concentration of Co2+ ions (Figure 3). Similarly, the size of osteoclasts was significantly increased by 2-fold in the presence of 100 μM of Co2+ as compared to unexposed PBMC control cultures (1662 ± 80 μm2 vs. 821 ± 75 μm2; p<0.0001). In contrast, 100 μM of Cr3+ had little effect on the number of nuclei per osteoclast or the size of newly formed osteoclasts. |
| |
|
|
Figure 3: Extent of human osteoclast formation after cultures with Co and Cr ions. (A) Representative photographs of long term human PBMCs cultures in the presence of M-CSF/sRANKL alone or in the presence of 100 μM of Co2+or Cr3+ and expression of OSCAR and CT-R. (B) Number of newly-formed osteoclasts in the presence of metal ions. White bar = M-CSF/sRANKL alone, black bars=Co2+-treated cultures and grey bars=Cr3+-treated cultures. *: p<0.05 compared to PBMCs treated with M-CSF/sRANKL. |
|
| |
| Our results indicated that in response to high concentrations of Co2+ and Cr3+, the TRACP levels were significantly increased (Figure 3). As the number of newly-formed osteoclasts was affected by the presence of Co2+ and Cr3+, we thought to assess the amount of dentine resorbed per osteoclast to evaluate whether not only osteoclast formation but also osteoclast activity was affected by the presence of metal ions. Compared to untreated cultures, Co2+ and Cr3+ reduced significantly and dosedependently the extent of resorption per osteoclast and as such the activity of newly-formed osteoclasts (Figure 3). This reduction in the activity of osteoclast is accompanied by a decrease in the size of lacunar pits (data not shown). |
| |
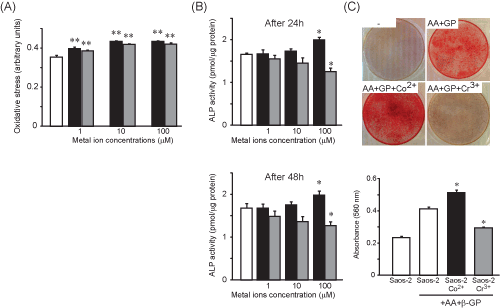
| Metal ions affect induce a rapid oxidative stress and affect the activity of alkaline phosphatase and the mineralisation capacity of SaOs-2 cells |
| |
| A rapid oxidative stress was noted in the presence of Co2+ and Cr3+ at all concentrations investigated (Figure 4A). After 24 h and 48 h cultures with high concentration of Co2+, the ALP activity was significantly increased by 1.2–fold and 1.2–fold respectively compared to untreated cells (Figure 4B). On the other hand, after 24 h and 48 h exposure to Cr3+ the ALP activity was significantly decreased by 1.3– fold and 1.3–fold respectively (Figure 4B). |
| |
|
|
Figure 4: Characterisation of newly-formed osteoclasts. White bar = M-CSF/sRANKL alone, black bars=Co2+- treated cultures and grey bars=Cr3+- treated cultures. *:
p<0.05 compared to PBMCs treated with M-CSF/sRANKL. |
|
| |
|
|
Figure 5: Oxidative stress (A), Alkaline phosphatase activity (B) and mineralisation rate (C) after cultures of human osteoblasts with Co2+ and Cr3+. White bar = untreated cells, black bars=Co2+-treated cultures and grey bars=Cr3+-treated cultures. *: p<0.05 compared to Saos-2 cultured in the absence of metal ions. |
|
| |
| Furthermore, cobalt increased significantly the mineralisation of the newly-formed matrix (Figure 4C) and indeed the mineralisation was increased by 1.25–fold compared with SaOs-2 cells cultured in the absence of metal ions (Figure 4C). On the other hand, chromium ions significantly decreased the mineralisation process (Figure 4C) and the extent of matrix mineralisation was clearly decreased by 1.4–fold compared with untreated SaOS-2 cells (Figure 4C). |
| |
| Discussion |
| |
| The use of metallic devices in orthopaedic has been of a major interest in reconstructive surgery due to their biomechanical properties and relatively good resistance to corrosion. However, depending on the metal morphology, some biological responses are encountered with metals. For example, titanium and vanadium are relatively biocompatible in bulk but very aggressive and toxic when in particles [18]. As body fluid is comprised of electrolytes, metals can also undergo corrosion and release metal ions in its surroundings [19]. Metals ions can directly affect the metabolism of bone cells. For example, gold ions are used to treat patient suffering with rheumatoid arthritis by inhibiting bone resorption and the release of inflammatory cytokines [20]. High concentrations of cobalt and chromium ions in serum and synovial fluid from patients with MoM implants have been previously noted [21,22]. However, the consequences of long-term exposure of circulating mononuclear cells and osteoblasts to high concentrations of Co and Cr ions remain unclear. In the present study, we have shown that Co2+ and Cr3+ did not significantly affect the viability of osteoclast precursors and osteoblasts. Co2+ was capable of increasing osteoclast formation from circulating precursors, but the activity of each individual osteoclast was reduced compared with untreated cells. On the other hand, Co2+ enhanced the activity of ALP in osteoblasts and contributed to increase the extent of matrix mineralisation. Cr3+ exhibited almost opposite effects. Chromium ions decreased osteoclast formation and osteoclast activity. A reduction of ALP activity and extent of mineralisation was also observed with chromium ions. |
| |
| Several studies reported a toxic effect of metal ions on osteoblasts and macrophages in vitro [11,12,23-25], however, in the present study; we did not find any significant evidence that Co2+ and Cr3+ induce human osteoblast or human circulating osteoclast precursor death. These discrepancies with previous published worked could be related to (i) the cells used in these studies which were mostly rodent cells or the MG-63 human osteosarcoma cell line and (ii) the dose of metal ions to which the cells were exposed to. In our study, the metal concentration was ranged from 1 μM to 100 μM and corresponded to levels of metal ions that are found in the synovial tissue of patients with MoM implants [3]. On the other hand, previous published reports used concentrations of metal ions ranged from 160 μM to 370 μM for Co2+ and from 480 μM to 2.8 mM for Cr3+. These concentrations are much higher than the concentrations we have used in our study and it is plausible that at these concentrations metal ions can induce cell death. |
| |
| In this study, we also reported that high concentration of Co2+, but not Cr3+, affected the rapid release of pro-inflammatory cytokine and the morphology of the newly-formed osteoclasts. The size of the cells and the number of nuclei per osteoclast were increased, indicating that more osteoclast precursors fused together. However, we did not investigate the molecular mechanism by which osteoclast precursor fusion is increased in the presence of high concentration of Co2+. One hypothesis could be that Co2+ increased the proliferation rate of osteoclast precursors and as such the increase in precursors could contribute to a greater rate of fusion and formation, but this hypothesis needs to be investigated in the future. Intracellular levels of TRAP were also increased in these cells in the presence of high concentrations of Co2+ and Cr3+. Helfrich et al., have recently shown that in some models of osteopetrosis, an increase in the intracellular levels of TRAP are correlated with an impairment of the release of the enzyme outside the cells to participate to the bone matrix degradation [26,27]. Indeed, in our study, we observed that osteoclast activity was reduced dose-dependently in the presence of Co2+ and Cr3+. This finding was surprising for Co2+ as the number of osteoclasts was increased; we expected that the activity would be also increased. Weinstein et al. reported similar findings in patients treated for prolonged period with alendronate [28]. These patients exhibited a significant increase in the number of osteoclasts compared with patients under placebo administration. However, the extent of osteoclastic bone resorption was decreased in alendronate-treated patients. However, the mechanism by which Co2+ reduced the activity of the newly-formed osteoclasts is unclear and will need further investigation. Nevertheless, it appears from our study that the effect of metal ions (both Co and Cr) on human osteoclasts is to decrease osteoclast activity and as such bone resorption. Recently, Andrews et al. reported similar findings on human osteoclasts, however, unlike our study, these authors did not characterise osteoclast parameters as size, number of nuclei and TRAP content [29]. Previous studies conducted on rodent osteoclasts have reported similar findings [9,10]. This is also in the agreement with the fact that periprosthetic osteolysis is rare with MoM implants. |
| |
| Metal ions at a concentration of 100 μM were also capable of affecting the osteoblastic response. Although Co2+ increased ALP activity and the mineralisation rate, Cr3+ decreased these two parameters. These results suggest that cobalt as an anabolic effect on osteoblast and matrix/ mineralisation formation whereas chromium (III) has a catabolic effect on matrix/mineralisation formation. Previous published studies reported that ALP activity was decreased in the rat FFC cell line after treatment with Cr3+12]. Anissian et al., reported that Co2+ ions induced a decrease in osteocalcin and collagen type I production [11], but the dose required in that study to see these effects was 1700-fold higher (170 mM) than the one we used (100 μM). It is likely that at a dose of 170 mM the effects of Co2+ are totally different than at a lower dose and hence could explain the discrepancy between that study and our study |
| |
| Chromium exists at different state of oxidation (Cr6+, Cr3+). One of the limitations of the present study is that we did not investigate the effects of Cr6+ on human osteoclast differentiation and osteoblast physiology. However, Cr6+ is an extremely reactive species and it is rapidly converted into a more stable stage, Cr3+ which persists in the tissue. |
| |
| Although osteolysis had been reported in the early failure of MoM prosthesis, its occurrence in patients with MoM implants is lower than in patients with polyethylene implants. Our results indicated that osteoclast activity was inhibited by the presence of Co or Cr ions and seems in opposition to osteolysis. However, MoM implants undergo degradation not only by corrosion, leading to the generation of metal ions, but also by friction of the articular surface leading to the release of metal nanoparticles. It is plausible that metal nanoparticles have a total different effect on the metabolism of both osteoclasts and osteoblasts and could influence osteolysis, but further investigations are required to validate this hypothesis. |
| |
| Conclusion |
| |
| In conclusion, Co and Cr ions directly acted on the physiology of human bone cells. Co and Cr ions decreased human osteoclast activity. On the other hand, metal ions had opposite effects on human osteoblast physiology, where Co2+ increased the mineralisation rate whereas Cr3+ decreased these two parameters. |
| |
| Acknowledgments |
| |
| The authors would like to acknowledge the funding from the Furlong Research Charitable Trust and the NIHR Biomedical Research Unit into Musculoskeletal Disease, the Nuffield Orthopaedic Centre and The University of Oxford. None of the author’s professional or financial affiliations have biased this presentation. |
| |
| |
| References |
| |
- Brodner W, Bitzan P, Meisinger V, Kaider A, Gottsauner-Wolf F, et al. (1997) Elevated serum cobalt with metal-on-metal articulating surfaces. J Bone Joint Surg Br 79: 316-321.
- Jacobs JJ, Skipor AK, Doorn PF, Campbell P, Schmalzried TP, et al. (1996) Cobalt and chromium concentrations in patients with metal on metal total hip replacements. Clin Orthop Relat Res S256-S263.
- De Smet K, De Haan R, Calistri A, Campbell PA, Ebramzadeh E, et al. (2008) Metal ion measurement as a diagnostic tool to identify problems with metal-on-metal hip resurfacing. J Bone Joint Surg 4: 202-208.
- Stulberg SD, Dorr LD, Freeman MA, Hungerford DS, Scott RD, et al. (1994) Knee challenges: what would you do? Orthopedics 17: 869-874.
- Mabilleau G, Filmon R, Petrov PK, Basle MF, Sabokbar A, et al. (2009) Cobalt, chromium and nickel affect hydroxyapatite crystal growth in vitro. Acta Biomater 6: 1556-1560.
- Beaule PE, Campbell P, Mirra J, Hooper JC, Schmalzried TP (2001) Osteolysis in a cementless, second generation metal-on-metal hip replacement. Clin Orthop Relat Res 159-165.
- McMinn D and Daniel J (2006) History and modern concepts in surface replacement. Proc Inst Mech Eng H 220: 239-251.
- Park YS, Moon YW, Lim SJ, Yang JM, Ahn G, et al. (2005) Early osteolysis following second-generation metal-on-metal hip replacement. J Bone Joint Surg Am 87: 1515-1521.
- Nichols KG and Puleo DA (1997) Effect of metal ions on the formation and function of osteoclastic cells in vitro. J Biomed Mater Res 35: 265-271.
- Rousselle AV, Heymann D, Demais V, Charrier C, Passuti N, et al. (2002) Influence of metal ion solutions on rabbit osteoclast activities in vitro. Histol Histopathol 17: 1025-1032.
- Anissian L, Stark A, Dahistrand H, Granberg B, Good V, et al. (2002) Cobalt ions influence proliferation and function of human osteoblast-like cells. Acta Orthop Scand 73: 369-374.
- McKay GC, Macnair R, MacDonald C, Grant MH (1996) Interactions of orthopaedic metals with an immortalized rat osteoblast cell line. Biomaterials 17: 1339-1344.
- Mabilleau G, Petrova NL, Edmonds ME, Sabokbar A (2008) Increased osteoclastic activity in acute Charcot's osteoarthopathy: the role of receptor activator of nuclear factor-kappaB ligand. Diabetologia 51: 1035-1040.
- Dumas A, Gaudin-Audrain C, Mabilleau G, Massin P, Hubert L, et al. (2006) The influence of processes for the purification of human bone allografts on the matrix surface and cytocompatibility. Biomaterials 27: 4204-4211.
- Mabilleau G and Sabokbar A (2009) Interleukin-32 promotes osteoclast differentiation but not osteoclast activation. PLoS One 4: e4173.
- Mabilleau G, Moreau MF, Filmon R, Basle MF, Chappard D (2004) Biodegradability of poly (2-hydroxyethyl methacrylate) in the presence of the J774.2 macrophage cell line. Biomaterials 25: 5155-5162.
- Mahoney DJ, Mikecz K, Ali T, Mabilleau G, Benayahu D, et al. (2008) TSG-6 regulates bone remodeling through inhibition of osteoblastogenesis and osteoclast activation. J Biol Chem 283: 25952-25962.
- Steinemann SG (1996) Metal implants and surface reactions. Injury 27 Suppl 3: SC16-SC22.
- Mabilleau G, Bourdon S, Joly-Guillou ML, Filmon R, Basle MF, et al. (2006) Influence of fluoride, hydrogen peroxide and lactic acid on the corrosion resistance of commercially pure titanium. Acta Biomater 2: 121-129.
- Lehman AJ, Esdaile JM, Klinkhoff AV, Grant E, Fitzgerald A, et al. (2005) A 48-week, randomized, double-blind, double-observer, placebo-controlled multicenter trial of combination methotrexate and intramuscular gold therapy in rheumatoid arthritis: results of the METGO study. Arthritis Rheum 52: 1360-1370.
- Grubl A, Marker M, Brodner W, Giurea A, Heinze G, et al. (2007) Long-term follow-up of metal-on-metal total hip replacement. J Orthop Res 25: 841-848.
- Korovessis P, Petsinis G, Repanti M, Repantis T (2006) Metallosis after contemporary metal- on-metal total hip arthroplasty. Five to nine-year follow-up. J Bone Joint Surg Am 88: 1183-1191.
- Catelas I, Petit A, Vali H, Fragiskatos C, Meilleur R, et al. (2005) Quantitative analysis of macrophage apoptosis vs. necrosis induced by cobalt and chromium ions in vitro. Biomaterials 26: 2441-2453.
- Fleury C, Petit A, Mwale F, Antoniou J, Zukor DJ, et al. (2006) Effect of cobalt and chromium ions on human MG-63 osteoblasts in vitro: morphology, cytotoxicity, and oxidative stress. Biomaterials 27: 3351-3360.
- Petit A, Mwale F, Tkaczyk C, Antoniou J, Zukor DJ, et al. (2006) Cobalt and chromium ions induce nitration of proteins in human U937 macrophages in vitro. J Biomed Mater Res A 79: 599-605.
- Helfrich M, Villa A, Frattini A, van Wesenbeeck L, van Hul We, et al. (2007) In vitro study of osteoclast phenotypes in different types of human osteopetrosis. American Bone and Mineral Society annual meeting.
- Reinholt FP, Hultenby K, Heinegård D, Marks SC Jr, Norgård M, et al. (1999) Extensive clear zone and defective ruffled border formation in osteoclasts of osteopetrotic (ia/ia) rats: implications for secretory function. Exp Cell Res 251: 477-491.
- Roos JC, Cox TM (2009) Giant osteoclast formation and long-term oral bisphosphonate therapy. N Engl J Med 360: 1676-1677.
- Andrews RE, Shah KM, Wilkinson JM, Gartland A (2011) Effects of cobalt and chromium ions at clinically equivalent concentrations after metal-on-metal hip replacement on human osteoblasts and osteoclasts: Implications for skeletal health. Bone 49: 717-723.
|
| |
| |