| Research Article |
Open Access |
|
| Masayoshi Yamaguchi |
| Department of Veterinary Pathobiology, The University of Missouri, Columbia, MO 65211, USA |
| *Corresponding authors: |
Masayoshi Yamaguchi
Department of Veterinary Pathobiology
The University of Missouri
205 Connaway Hall, 1600 East
Rollins Street, Columbia
MO 65211, USA
E-mail: yamamasa1155@yahoo.co.jp |
|
| |
| Received June 20, 2012; Published July 30, 2012 |
| |
| Citation: Yamaguchi M (2012) Bone Anabolic Effect of Wasbi Leafstalk Component: Its Related Phytochemical p-hydroxycinnamic Acid Prevents Osteoporosis. 1: 194. doi:10.4172/scientificreports.194 |
| |
| Copyright: © 2012 Yamaguchi M. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. |
| |
| Abstract |
| |
| Bone homeostasis is maintained through a balance between osteoblastic bone formation and osteoclastic bone resorption. Aging induces bone loss due to decreased osteoblastic bone formation and increased osteoclastic bone resorption. Osteoporosis with its accompanying decrease in bone mass is widely recognized as a major public heath problem. Pharmacologic and nutritional factors may play a role in the prevention and treatment of bone loss with aging. Among various food and plants tested, the active component to stimulate bone mineralization in mouse bone tissues in vitro was found to be present in the leafstalk of wasabi (Wasabi japanica MATSUM). The material with a low molecular weight of 158 may be the active component in wasabi leafstalk that stimulates bone mineralization. When rat femoral tissues were cultured in a medium containing cinnamic acid, p-hydroxycinnamic acid (HCA), ferulic acid, caffeic acid, or 3,4-dimethoxycinnamic acid that is found in wasabi leafstalk extracts, HCA was uniquely found to have a potential-anabolic effect on bone mineralization in vitro. HCA was found to stimulate osteoblastic bone formation and suppresses osteoclastic bone resorption in vitro. The oral administration of HCA was found to have a restorative effect on bone loss induced with ovarietomy and diabetic state. Thus, HCA may have a role in the prevention and treatment of osteoporosis. |
| |
| Keywords |
| |
| Wasabi leafstalk; p-Hydroxycinnamic acid; Osteoblastic bone formation; Osteoclastic bone resorption; Osteoporosis; Diabetes |
| |
| Introduction |
| |
| Bone homeostasis is maintained through a delicate balance between osteoblastic bone formation and osteoclastic bone resorption. Numerous pathological processes have the capacity to disrupt this equilibrium leading to conditions where the rate of bone resorption outpaces the rate of bone formation leading to osteoporosis, a devastating bone disease that is widely recognized as a major public health threat [1]. Postmenopausal osteoporosis, a consequence of ovarian hormone deficiency, is the archetypal osteoporotic condition in women after menopause and leads to bone destruction though complex and diverse metabolic and biochemical changes [1]. Osteoporosis is induced with decrease in bone mass. It is widely recognized as a major public health problem. The most dramatic expression of the disease is represented by fractures of the proximal femur for which the number increases as the population ages [2]. |
| |
| Diets and nutritional factors may have the potential effect to delay degenerative bone disorders such as osteoporosis. There is growing evidence that the supplementation of nutritional and food factors may have the preventive effect on bone loss that is induced in animal model of osteoporosis and in human subjects [3-6]. Chemical compounds in food and plants, which regulate bone homeostasis, have been to be worthy of notice in maintaining of bone health and prevention of bone loss with increasing age [7-12]. It appears increasingly likely that as-yet-unidentified factors found in the daily consumption of fruits or vegetables may play a role in the building of optimal peak bone mass and in the preservation of decreased bone mass with aging. |
| |
| It is found that, among various fruits, plants and vegetables, wasabi leafstalk has a potential-anabolic effect on bone and that its active component may be identified as p-hydroxycinnamic acid (HCA). This phenolic acid has been shown to have osteoblastic bone formation and osteoclastogenic bone resorption in vitro. HCA has been found to have a preventive effect on bone loss with patho-physiological states including ovariectomy and diabetes state. |
| |
| This review has been written to outline the recent advances concerning the role of HCA in the regulation of bone homeostasis and in the prevention of osteoporosis. |
| |
| Cellular Regulation of Bone Homeostasis |
| |
| Bone metabolism plays a physiological role to maintain skeletal structure and to regulate mineral homeostasis. Bone homeostasis is regulated by the functions of osteoblasts, osteoclasts, and osteocytes that are major cells in bone tissues. Bone remodeling and modeling underpin development and maintenance of the skeletal system [13,14]. |
| |
| Bone modeling is responsible for growth and mechanically induced adaptation of bone and requires that the processes of bone formation and bone removal (resorption). Bone remodeling is responsible for removal and repair of damaged bone to maintain integrity of the adult skeleton and mineral homeostasis [14]. This tightly coordinated event requires the synchronized activities of multiple cellular participants to ensure that bone resorption and formation occur sequentially at the same anatomical location to preserve bone mass [24]. In addition to the traditional bone cells (osteoclasts, osteoblasts, and osteocytes) that are necessary for bone remodeling, several immune cells have also been implicated in bone disease [14]. |
| |
| Osteoclasts, which develop from hematopoietic progenitors, are uniquely adapted to remove mineralized bone matrix. A resorptive stimulus firstly triggers recruitment of osteoclasts to a site on the bone surface. This is followed by active resorption by osteoclasts, after which cells withdraw from the bone surface and mononuclear phagocytic cells appear on the newly resorbed surface. Osteoblasts arising from local mesenchymal stem cells assemble at the bottom of the cavity, and bone formation begins. After the resorbed lacunar pit is filled with new osteoid, osteoblasts become flatter and less active with the final newly remodeled bone surface lined by flat lining cells. As bone formation progresses, some osteoblasts are entombed within the matrix as osteocytes but the majority dies by apoptosis. This event appears to act as a beacon for osteoclast recruitment and generation of a new basic multicellular unit. |
| |
| Bone is major storage site for growth factors [15-17]. Growth factors, which are produced by osteoblasts, diffuse into newly deposited osteoid and are stored in the bone matrx including isulin-like growth factors (IGF- I and II), transforming growth factor-β1 (TGF-β1), platelet-derived growth factor (PDGF), and bone morphologic protein (BMP). These bone-derived factors, which can be liberated during subsequent periods of bone resorption, act in an autocrine, paracrine, or delayed paracrine fashion in the local microenvironment of the bone surface. |
| |
| The process of bone remodeling that makes bone unique among organs and tissues, and add so many levels of complexity with respect to interactions along the remodeling sequence by systemic influences (hormones), stress action (physical activity/weight bearing), growth factors and cytokines produced by the bone cells or factors that come from nearby cells in the marrow tissues. |
| |
| Bone Anabolic Factor in Wasabi Leafstalk Extracts |
| |
| The effects of various foods and plants on bone tissues have been poorly understood. Then, the effects of 20% ethanol solution extracts obtained from various foods and plants on bone calcium content is examined using mouse bone tissues in vitro. Among various food and plants (including shiitake, loquat leaf, cherry leaf, gabaron tea, green tea, muskmelon, tomato, blueberry, and wasabi leafstalk) used, the extracts of wasabi leafstalk (Wasabi japanica MATSUM) were found to have a potential-anabolic effect on bone calcification in mouse calvaria tissue culture in vitro [18]. This was the first time finding. |
| |
| Stimulatory Effect of Wasabi Leafstalk Extract on Bone Mineralization In vitro |
| |
| The presence of wasabi leafstalk extract (10 μg/ml of culture medium), which wasabi leafstalk extract was obtained from a homogenate with 20% ethanol, caused a significant increase in calcium content and alkaline phosphatase activity, which participates in mineralization in the bone tissues [19,20]. The wasabi leafstalk extractinduced increase in bone calcium content was completely depressed in the presence of cycloheximide, an inhibitor of protein synthesis, suggesting that the effect of wasabi leafstalk extracts is resulted from newly synthesized protein components [19]. In addition, wasabi leafstalk extract has been shown to have a suppressive effect on parathyroid hormone (PTH)-induced osteoclast-like cell formation in a mouse marrow culture system, suggesting that the component inhibits bone resorption [19]. PTH has been known to stimulate osteoclastogenesis and bone resorption [19]. |
| |
| The stimulatory effect of wasabi leafstalk extract on bone mineralization is compared with that of insulin, IGF-I, 17β-estradiol, and genistein [21]. The combination of 17β-estradiol (10-10 M) and wasabi leafstalk extract (15 μg/ml) had an additive effect on the increase in bone calcium content [21]. Also, the combination of either insulin (10-8 or 10-7 M) with wasabi leafstalk extract (15 μg/ml) had an additive effect the increase in bone calcium content [21]. This additive effect was not seen in the case of IGF-I (10-8 M). Wasabi leafstalk extract has an enhancing effect on the anabolic action of 17β-estradiol or insulin, which physiologically regulates bone formation in vitro [21]. |
| |
| The oral administration of wasabi leafstalk extract to young rats cased a significant increase in alkaline phosphatase activities, calcium and DNA contents in the femoral-metaphyseal tissues (trabecullar bone) [22]. In the femoral-diaphyseal tissues (cortical bone), alkaline phosphatase activity and calcium content were increased after administration of wasabi extract [22]. This administration had a greater effect on the metaphyseal tissues than on the diaphyseal tissues [22]. Many bone cells (including chondrocytes and osteoblasts) are localized in the metaphyseal tissues. The finding, that the administration of wasabi leafstalk extract produces an increase in DNA content in the metaphyseal tissues, suggests that the administration stimulates the proliferation of bone cells. The anabolic effect of wasabi leafstalk extract on bone tissues may be based on newly synthesized protein components [22]. |
| |
| Whether wasabi leafstalk extract has a preventive effect on bone loss with increasing age, which induces osteoporosis, is examined in vitro and in vivo [23]. Bone metabolism in the femoral-diaphyseal and metaphyseal tissues is deteriorated with increasing age (50 weeks old), as compared with that of young rats (4 weeks old) [24]. The presence of wasabi leafstalk extract in the culture medium caused a significant increase in alkaline phosphatase activity, calsium and DNA contents in the femoral-diaphyseal and metaphyseal tissues of aged female rats in vitro [23]. The effect of wasabi leafstalk extract in increasing these bone components was completely depressed in the presence of an inhibitor of protein synthesis [23]. This finding suggests that the anabolic effect of wasabi leafstalk extract on bone is resulted from newly synthesized protein components in aged rat bone tissues. |
| |
| Moreover, the oral administration of wasabi leafstalk extract was found to induce an increase in alkaline phosphatase activity, calcium and DNA contents in the femoral-diaphyseal and metaphyseal tissues of aged female rats in vivo [23]. The administration of wasabi leafstalk extract did not cause any change in body weight, serum calcium or inorganic phosphorus concentrations in aged female rats, indicating that the extract may not have a toxic effect [23]. Thus, the intake of wasabi leafstalk extract may have a preventive effect on the deterioration of bone metabolism with increasing age. Presumably, dietary wasabi leafstalk has a preventive role in the progression of osteoporosis with aging. |
| |
| Active Component of Wasabi Leafstalk Extract |
| |
| |
| The components of wasabi leafstalk extracts that increase bone calcium content were stable when the components were treated with heat, acidity, or alkalinization [25]. The active components of wasabi leafstalk extract thus may not be a protein. |
| |
| The active component in wasabi leafstalk is purified [25]. Wasabi leafstalk extract was obtained from a homogenate with 20% ethanol. The active component, which was found in the ethanol extract, was purified upon gel filtration chromatography with a HiLoad 26/80 Superdex 30-pg column and reverse-phase chromatography on a Resource 15 RPC 3-ml column. The result of ESI mass spectra for the purified active component showed that the material had a molecular weight of 158 [25]. |
| |
| The material with a low molecular weight of 158 may be the active component in wasabi leafstalk that stimulates bone mineralization. Cinnamic acid, p-hydroxycinnamic acid (HCA), ferulic acid, caffeic acid, or 3,4-dimethoxycinnamic acid (DCA) has been found to be present in the wasabi leafstalk. When rat femoral tissues were cultured in a medium containing these phenolic acids, HCA was found to uniquely have a potential-anabolic effect on bone mineralization in vitro [26]. The molecular weight of HCA is 164. We judged that HCA was an active component in wasabi leafstalk that reveals bone anabolic effect. |
| |
| HCA Stimulates Osteoblastic Bone Formation in vitro |
| |
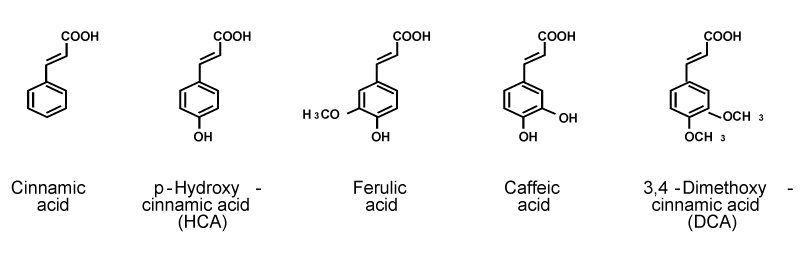
| Cinnamic acid is present in many plant and fruits. HCA is an intermediate-metabolic substance in plants and fruits, and it is synthesized from tyrosine. The effect of cinnamic acid or its related compounds, which their chemical structures are shown in Figure 1, on bone metabolism is examined in vitro using rat femoral tissues [25]. Among these phenolic acids, HCA has been found to uniquely have a potential-anabolic effect on bone tissues [25]. This was a first-time finding. |
| |
|
|
Figure 1: Chemical structures of cinnamic acid and its related compounds. The molecular weight of cinnamic acid, p-hydroxycinnamic acid (HCA), ferulic acid, caffeic acid, and 3,4-dimethoxycinnamic acid (DCA) is 148.2, 164.2, 194.2, 180.2, and 208.3, respectively. |
|
| |
| HCA has an Anabolic Effect on Bone Tissues |
| |
| The effect of cinnamic acid and its related compounds on bone mineralization is not determined so far. Then, the rat femoraldaiphyseal or metaphyseal tissues were cultured for 48 hours in a medium containing cinnamic acid, p-hydroxycinnamic acid (HCA), ferulic acid, caffeic acid, or 3,4-dimethoxycinnamic acid in vitro [26]. Culture with HCA (10-5 or 10-4 M) caused an increase in calcium content in the diaphyseal and metaphyseal tissues [26]. This effect was not observed after culture with cinnamic acid or other compounds at the concentration of 10-5 or 10-4 M. Also, alkaline phosphatase activity and DNA content in the diaphyseal or metaphyseal tissues was increased after culture with HCA (10-5 or 10-4 M) [26]. The effects of HCA (10-4 M) in increasing alkaline phosphatase activity, DNA and calcium contents in the diaphyseal or metaphyseal tissues were completely depressed in the presence of cycloheximide (10-6 M), an inhibitor of protein synthesis. Thus, HCA has been found to have a unique-anabolic effect on bone metabolism, which results from newly synthesized protein components [26]. |
| |
| DNA content in the bone tissues may be partly involved in the number of bone cells including osteoblasts, osteocytes, and osteoclasts. HCA increased DNA content in the femoral-diaphyseal and metaphyseal tissues of rats in vitro and stimulated bonemineralization. HCA may stimulate proliferation of osteoblastic cells in the bone tissues in vitro and has a stimulatory effect on bone formation. |
| |
| As the chemical structure of cinnamic acid and its related compounds is shown in Figure 1, HCA is hydroxylated at 4-position of cinnamic acid. This chemical form may have an anabolic effect on bone metabolism, suggesting a relationship of structure and activity of cinnamic acid. |
| |
| HCA Stimulates Osteoblastogenesis |
| |
| Osteoblastic MC3T3-E1 cells in vitro were cultured for 72 hours in a minimum essential medium containing 10% fetal bovine serum (FBS) and the subconfluent cells were changed to a medium containing either vehicle or HCA (10-7 to 10-5 M) without FBS [27]. Culture with HCA (10-7 to 10-5 M) did not have a significant effect on cell proliferation of osteoblastic cells in reaching subconfluent monolayers [27]. Also, the number of osteoblastic cells after reaching to subconfluent was not changed after culture with HCA, indicating that the compound does not induce cell death [26]. However, culture with HCA caused an increase in DNA content in osteoblastic cells [27]. The effect of HCA in increasing DNA content in osteoblastic cells was also seen in the absence of FBS [27]. HCA may have a stimulating effect on DNA synthesis in osteoblastic cells. |
| |
| Alkaline phosphatase is involved in mineralization in osteoblastic cells [20]. Alkaline phosphatase activity in osteoblastic cells was significantly increased after culture with HCA (10-7 to 10-5 M) for 24 - 72 hours [27], indicating that HCA stimulates cell differentiation in osteoblastic cells. The results of Alizarin red stain showed that the prolonged culture with HCA markedly stimulates mineralization in osteoblastic cells; the mineralization was stimulated after culture with HCA (10-8 to 10-5 M) for 7, 14, or 21 days [27]. This finding support the view that culture with HCA stimulates the differentiation of osteoblastic cells and that promotes mineralization in the cells. In addition, HCA caused a significant increase in calcium content in rat femoral tissue culture in vitro. Thus, HCA has been demonstrated to have a stimulatory effect on osteoblastic differentiation and mineralization [27]. |
| |
| HCA Suppresses Osteoclastic Bone Resorption in vitro |
| |
| Osteoclasts are generated from bone marrow stem cells. Receptor activator of the nuclear factor kappa B (NF-κB) (RANK) ligand (RANKL) plays a pivotal role in osteoclastogenesis from bone marrow cells [28,29]. RANKL is produced from osteoblasts in response to osteoporotic factors, such as PTH, PGE2, and 1,25-dihydroxyvitamin D3 (VD3) [28-30]. RANKL acts osteoclast progenitors and stimulates osteoclast differentiation [28,29]. Osteoclastic cells are differentiated from bone marrow stromal cells. Osteoclastogenesis is stimulated by macrophage colony-stimulating factor (M-CSF) and RANKL in vitro [28,29]. A soluble fragment containing part of the extra-cellular domain of RANKL (the carboxyterminal half of the protein, amino acids (158-316) is capable of promoting osteoclastogenesis in the presence of M-CSF [28, 29]. |
| |
| The receptor protein RANK for RANKL is expressed on the surface of osteoclast progenitors. The interaction of RANKL with its receptor RANK leads to the recruitment of the signaling adaptor molecules TRAFs (TNF receptor-associated factors) to the receptor complex and the activation of nuclear factor-κB (NF-κB) and c-Jun N-terminal kinase (JNK) [29,30]. The protein kinase C family enzyme has a role in the regulation of osteoclast formation and function potentially by participating in the extracellular signal-regulated kinase (ERK) signaling pathway of M-CSF and RANKL [29,30]. |
| |
| HCA has been found to have a suppressive effect on bone resorption induced by bone-resorbing factors in the bone tissue culture in vitro. PTH or VD3 is known as bone-resorbing factors [30,31]. Culture with PTH caused a decrease in calcium content and an increase in the activity of tartrate-resistant acid phosphatase (TRACP), which is a marker enzyme in osteoclastic cells, in the diaphyseal or metaphyseal tissues and a corresponding elevation in medium glucose consumption and lactic acid production by the bone tissues [26]. These alterations were completely depressed after culture with HCA (10-5 or 10-4 M). Thus, HCA has been shown to have a suppressive effect on bone resorption in the bone tissue culture in vitro [26]. |
| |
| Culture with HCA has also been found to have a suppressive effect on PTH-, prostaglandin E2 (PGE2)-, or tumor necrosis factor-α (TNF- α)-induced osteoclast-like cell formation from mouse bone marrow in vitro [32]. HCA did not have an effect on the proliferation of bone marrow cells, suggesting that the compound did not have a toxic effect on the cells [32]. The suppressive effect of HCA on osteoclast-like cell formation was remarkable at the earlier stage of the differentiation to osteoclasts in bone marrow cultures [32]. In addition, the suppressive effect of HCA was also observed at the later stage of osteoclastogenesis [32]. HCA may have an effect on the process of the differentiation from mononuclear osteoclast to osteoclast [32]. |
| |
| Moreover, HCA has been shown to suppress osteoclast-like cell formation induced by PTH or PGE2 [32]. The suppressive effect of HCA was partly involved in RANKL expression and/or RANKL action, which are related to the effect of PTH and PGE2. Culture with HCA suppressed RANKL plus M-CSF-induced osteoclast-like cell formation in mouse bone marrow culture in vitro [32]. Presumably, the suppressive effect of HCA is involved in RANKL expression and/or RANKL action, which are related to the effect of PTH or PGE2. |
| |
| TNF-α is an autocrine factor in osteoclasts, promoting their differentiation and mediating RANKL-induced osteoclastogenesis [33]. TNF-α has also been shown to mediate via its p55 receptor in lypoplysaccaride-stimulated osteoclastogenesis [33]. TNF-α-induced osteoclast-like cell formation in mouse bone marrow cultures was significantly suppressed after culture with HCA [32]. HCA may suppress osteoclastogenesis that is mediated through RANKL and TNF-α. |
| |
| Phorbol 12-myristate 13-acetate (PMA) is an activator of protein kinase C [34]. PMA stimulated osteoclast-like cell formation in mouse bone marrow cultures, and the PMA-induced osteoclastogenesis was suppressed after culture with HCA [32]. Moreover, HCA was found to have a suppressive effect on Dibutyryl cyclic AMP-induced osteoclastlike cell formation in mouse bone marrow cultures [32]. The activation of the protein kinase C or protein kinase A may be related to RANKL signaling in osteoclastogeneasis. HCA may have a suppressive effect on osteoclastogenesis that is mediated through signaling mechanim of protein kinase C or protein kinase A. |
| |
| Culture with HCA caused an inhibition of RANKL plus M-CSFinduced osteoclast-like cell formation in mouse bone marrow culture in vitro [32]. This observation may support the view that HCA represses the binding of RANKL to receptor RANK and that suppresses RANKL signaling in osteoclastogenesis. |
| |
| The effect of HCA in inhibiting RANKL plus M-CSF-induced osteoclastogenesis was completely depressed in the presence of cycloheximide, an inhibitor of protein synthesis in translational processes [32]. The stimulatory effect of RANKL plus M-CSF on osteoclastogenesis was not suppressed in the presence of cycloheximide or 5,6-dichloro-1-β-D-ribofuranosylbenzimidazole (DRB), an inhibitor of transcriptional processes [32]. It is possible that the suppressive effect of HCA on osteoclastogenesis is partly involved in newly synthesized protein components, which induce suppressor protein. Whether HCA has an effect on the expression of osteoprotegerin (OPG), a regulated suppressor of osteoclast differentiation [29,30] in osteoclasts, is unknown. |
| |
| HCA Stimulates Osteoblastogenesis and Suppresses Osteoclastogenesis through Suppression of NF-κB Activation |
| |
| The NF-κB signal transduction pathway has long been recognized as critical for osteoclast development and function [35,36] and double knockout of p50 and p52 NF-κB subunits leads to osteopetrosis due to a severe defect in osteoclast differentiation in these mice [37]. Furthermore, it has been reported that NF-κB signaling represses basal osteoblast differentiation and mineralization in MC3T3 cells and antagonizes TGF-β1 and BMP-2 mediated MC3T3 mineralization by downregulating Smad activation [38]. Other studies have found that NF-κB signaling antagonizes Smad activation in Saos2 osteosarcoma cells by a mechanism involving induction of inhibitory Smad7 [39]. |
| |
| TNF-α is an inflammatory cytokine that antagonizes bone formation in vivo and osteoblastic differentiation in vitro [38,39]. These effects are mediated in large measure through NF-B signaling. Inflammatory levels of TNF-α are known to impact bone formation; however, it has been recently reported that basal TNF-α levels in vivo also dramatically lower the basal bone formation rate [38]. HCA may promote accumulation of basal bone mass, and forestall bone loss during osteoporotic states, in part by antagonizing TNF-α-induced NF-κB. Interestingly, HCA prevented TNF-α-induced suppression of mineralization in osteoblastic 3T3-E1 cells and repressed TNF-α- induced NF-κB activation in the osteoblastic cells [40,41]. Culture with HCA was also found to suppress the differentiation of preosteoclast (RAW267.4 cells) to osteoclasts induced by RANKL through suppression of NF-κB activation [41]. |
| |
| The 5 members of the mammalian NF-κB family RelA/p65, RelB, c-Rel, NF-κB1/p50, and NF-κB2/p52 are activated through one of two specific pathways, the canonical NF-κB pathway or an alternative pathway [42]. In the canonical pathway activation of the inhibitor of IκB kinase (IKK) complex, leads to phosphorylation of the NF- κB-associated IκBα, catalyzing its ubiquitination and proteasomal degradation, and in the process releasing active NF-κB dimmers that translocate to the nucleus and enhance transcription of target genes [42]. In the alternative NF-κB pathway, NF-κB-inducing kinase (NIK) and IKKα target p100 for proteolytic processing, thereby releasing active RelB-containing dimmers [42]. Differential regulation of these two pathways by HCA could provide a more detail explanation for the suppressive effect observed in TNFα-induced NF-κB activation. |
| |
| Cellular mechanism by which HCA stimulates osteoblastic bone formation and suppresses osteoclastic bone resorption is mediated through suppression of NF-κB activation, as shown in Figure 2. |
| |
|
|
Figure 2: p-Hydroxycinnamic acid (HCA) reveals anabolic effect on bone. HCA stimulates osteoblastic bone formation and suppresses osteoclastic bone resorption, thereby increasing bone mass. The osteogenic effect of HCA is mediated through suppression of NF-κB activation. |
|
| |
| HCA Prevents Osteoporosis In vivo |
| |
| Anabolic effect of HCA on normal bone in vivo |
| |
| The anabolic effects of HCA on the bone of rats in vivo are shown [43-45]. Rats were orally administered HCA (10, 20, or 50 mg/ kg body weight) once daily for seven days [43]. The administration of HCA did not cause a significant change in body weight or serum calcium and inorganic phosphorus levels. Alkaline phosphatase activity, DNA and calcium contents in the diaphyseal and metaphyseal tissues were increased after the administration of HCA (20 or 50 mg/kg) [43]. Diaphyseal calcium and metaphyseal DNA contents were increased with the dose of HCA 10 mg/kg. These findings suggest that the oral intake of HCA induces anabolic effects on bone mineralization in normal growing rats. |
| |
| The activity of TRACP, which is a marker enzyme of osteoclastic bone resorption, is enhanced by bone-resorbing factors [46]. The oral administration of HCA (10, 20, or 50 mg/kg) caused a significant decrease in TRACP activity in the femoral-diaphyseal and – metaphyseal tissues of rats [43]. This suggests that the administration of HCA induces a decrease in bone-resorbing activity in the femoral tissues of rats in vivo. |
| |
| The mixture of phenolic acids found in the serum of young rats fed blueberries has been reported to stimulate osteoblast differentiation, resulting in significantly increased bone mass [47]. Greater bone formation in blueberries diet-fed animals is associated with increase in osteoblast progenitors and osteoblast differentiation and reduced osteoclastogenesis [47]. Most of the phenolic acids in the circulation of blueberries diet-fed animals are either metabolites or breakdown products of polyphenols and phenolic acids found in blueberries [47]. Interestingly, HCA is found in the serum after diet of blueberries [47]. |
| |
| HCA Prevents Ovariectomy-Induced Bone Loss |
| |
| The preventive effect of HCA on osteoporosis is examined using ovariectomized (OVX) rats, a animal model for osteoporosis [44]. HCA (0.25 or 0.5 mg/kg body weight) was orally administered once daily for 30 days to OVX rats. The analysis using a peripheral quantitative computed tomography (pQCT) showed that OVX caused bone loss in the femoral-metaphyseal tissues [43]. This bone loss was restored after the administration of HCA (2.5 or 5 mg/kg body weight) to OVX rats [44]. Mineral content, mineral density, and polar strength strain index in the femoral-metaphyseal tissues were decreased in OVX rats [44]. These decreases were restored after the administration of HCA (5 mg/kg) to OVX rats [44]. Especially, the polar stress strain index is an indicator of bone strength. This observation suggests that the oral administration of HCA causes a functional change in the bone of OVX rats. |
| |
| Moreover, OVX caused a decrease in calcium content or alkaline phosphatase activity in the femoral-diaphyseal and -metaphyseal tissues. These decreases were also restored after the administration of HCA (2.5 or 5 mg/kg) to OVX rats [44]. |
| |
| Thus, HCA has been shown to have preventive effects on OVXinduced bone loss of rats in vivo. Presumably, HCA stimulates osteoblastic bone formation and inhibits osteoclastic bone resorption in vivo, thereby increasing bone mass in OVX rats. |
| |
| The dose with HCA 2.5 or 5 mg/kg body weight had a preventive effect on the decrease in alakaline phosphatase activity and calcium content or the increase in DNA content in the femoral-diaphyseal tissues of OVX rats, although the dose of HCA 5 mg/kg had great potential effects in the restoration of OVX-induced bone change [44]. This suggests that the intake of HCA less than 2.5 mg/day/kg body weight has preventive effects on bone loss. The amount of intake of HCA in fruit and vegetables is unknown. However, the dietary intake of phytocomponent HCA may have a role in the prevention of bone loss with aging. HCA may have a preventive effect on bone loss with increasing age. |
| |
| HCA has Anti-Diabetic Effects In vivo |
| |
| Bone loss is induced in the diabetic state [46]. Streptozotocin (STZ) induces type I diabetes. The preventive effect of HCA on bone loss induced in STZ-diabetic rats is examined in vivo [44]. Rats received a single subcutaneous administration of STZ (60 mg/kg body weight), and then the animals were orally administered HCA (2.5, 5, or 10 mg/ kg body weight) once daily for 14 days. STZ administration caused a decrease in body weight and a significant increase in serum glucose, triglyceride, and calcium levels, indicating a diabetic state [44]. These alterations were prevented after the administration of HCA (2.5, 5, or 10 mg/kg) [44]. This was a novel finding. The oral intake of HCA has a restorative effect on serum biochemical findings that are involved in diabetes in vivo. HCA may have a role in the prevention of diabetic states. |
| |
| Serum calcium concentration was found to increase in STZ-diabetic rats [44]. Intestinal calcium absorption has been shown to be impaired in the diabetic state [48]. The increase in serum calcium concentration in STZ-diabetic rats may result from the release of calcium from the bone tissues; the femoral calcium content was found to markedly decrease in STZ-diabetic rats [48]. The oral administration of HCA to the diabetic rats had a significant preventive effect on hypercalcemia and bone calcium loss in the diabetic state. Thus, the intake of HCA may have a preventive effect on bone resorption in diabetic rats. Calcium content in the femoral-diaphyseal and -metaphyseal tissues was decreased in STZ-diabetic rats [44]. Also, this decrease was prevented after administration of HCA (2.5, 5, or 10 mg/kg) [44]. The intake of dietary HCA may have a restorative effect on bone loss in diabetic state. Alkaline phosphatase activity in the diaphyseal and metaphyseal tissues was decreased in STZ-diabetic rats [44]. The decrease in diaphyseal alkaline phosphatase activity in STZ-diabetic rats was restored after administration of HCA (5 and 10 mg/kg). This enzyme participates in osteoblastic mineralization. The femoral alkaline phosphatase activity was decreased in diabetic rats, suggesting that osteoblastic bone mineralization is impaired in the diabetic state. |
| |
| In addition, the diaphyseal DNA content was also decreased in STZ-diabetic rats. Administration of HCA (2.5, 5, or 10 mg/kg) caused an increase in DNA content in the diaphyseal and metaphyseal tissues in STZ-diabetic rats. DNA content in the bone tissues is an index of the number of existing bone cells [15]. HCA may stimulate an increase in bone cells including osteoblastic cells in the diaphyseal and metaphyseal tissues of STZ-diabetic rats in vivo. This may be partly contributed to the increase in bone calcium content in STZ-diabetic rats. Presumably, the intake of HCA has a stimulatory effect on osteoblastic bone formation in diabetic state. |
| |
| Thus, the intake of HCA has a preventive effect on bone loss in STZ-diabetic rats and has a restorative effect on serum biochemical findings in the diabetic state, suggesting that HCA has a restorative effect on diabetic state. HCA may have a role in the prevention and treatment of osteoporosis in diabetic state. |
| |
| As mentioned above, HCA has been demonstrated to have an anabolic effect on bone mass due to stimulating osteoblastic bone formation and inhibiting osteoclastic bone resorption and has a restorative effect on bone loss induced in OVX and diabetic rats. The supplemental intake of HCA may have a role in the prevention and treatment of osteoporosis. |
| |
| Summary and Prospect |
| |
| Diet and nutritional status are critical factors that influence bone development and aged bone loss. Phytochemicals as food functional factor may play a role in delay degenerative bone disorders with aging and in treatment of osteoporosis with various pathophysiological conditions. Phytocomponent p-hydroxycinnamic acid (HCA) was found to have the osteogenic effects due to stimulating osteoblastic bone formation and suppressing osteoclastic bone resorption in vitro and the preventive effects on bone loss, which is induced with ovariectomy and diabetes in vivo. |
| |
| The finding, that HCA has an osteogenic effect, was the first time in this study. Molecular mechanism by which HCA stimulates osteogenesis and suppresses osteoclastogenesis may be mediated through suppression of NF-κB activation. NF-κB is a signaling factor that plays a pivotal role in the regulation of bone homeostasis regulated through inflammatory cytokines. Whether HCA has an effect on other transcription and signaling systems, however, remains to be elucidated. |
| |
| HCA had a potently stimulating effect on osteogenesis as compared with that of other phenolic acids (including ferulic acid, caffeic acid, or 3,4-dimethoxycinnamic acid), indicating a relationship with chemical structure and osteogenic activity. Phytochemical HCA may be usefulness as pharmacologic tool to treat osteoporosis. HCA analog with more potently effect may be developed. Clinical studies in bone disorder are expected through further experiments. |
| |
| |
| References |
| |
- Weitzmann MN, Pacifici R (2006) Estrogen deficiency and bone loss: an inflammatory tale. J Clin Invest 116: 1186-1194.
- Johnell O, Kanis JA (2006) An estimate of the worldwide prevalence and disability associated with osteoporotic fractures. Osteoporos Int 17: 1726-1733.
- Bonjour J-P, Schurch M-A, Rizzori R (1996) Nutritional aspects of hip fracture. Bone 18: 1395-1445
- Yamaguchi M (2002) Isoflavone and bone metabolism: Its cellular mechanism and preventive role in bone loss. J Health Sci 48: 209-222
- Yamaguchi M (2006) Regulatory mechanism of food factors in bone metabolism and prevention of osteoporosis. Yakugaku Zasshi 126: 1117-1137.
- Yamaguchi M (2010) Role of nutritional zinc in the prevention of osteoporosis. Mol Cell Biochem 338: 241-254.
- Yamaguchi M, Hamamoto R, Uchiyama S, Ishiyama K (2007) Effects of flavonoid on calcium content in femoral tissue culture and parathyroid hormone-stimulated osteoclastogenesis in bone marrow culture in vitro. Mol Cell Biochem 303: 83-88.
- Yamaguchi M, Weitzmann MN (2011) Vitamin K2 stimulates osteoblastogenesis and suppresses osteoclastogenesis by NF-?B activation. Int J Mol Med 27:3-14
- Uchiyama S, Yamaguchi M (2002) Anabolic effect of marine alga Sargassum horneri extract on bone components in the femoral-diaphyseal and -metaphyseal tissues of young and aged rats in vivo. J Health Sci 48: 325-330
- Yamaguchi M, Uchiyama S, Nakagawa T (2007) Preventive effects ofbee pollen Cistus ladaniferus extract on bone loss in ovariectomiaed rats in vivo. J Health Sci 53: 571-575
- Yamaguchi M (2008) ß-Cryptoxanthin and bone metabolism: The preventive role in osteoporosis. J Health Sci 54: 356-369
- Yamaguchi M (2010) In: Nutritional Factors and Osteoporosis
- Prevention. Nova Science Publishers, Inc., New York, 2010
- Parfitt AM (1990) Bone-forming cells in clinical conditions. In: Hall, B. K., editor. Bone Volume 1. The Osteoblast and Osteocyte. Boca Raton, FL: Telford Press and CRC Press p351-429
- Raggatt LJ, Partridge NC (2010) Cellular and molecular mechanisms of bone remodeling. J Biol Chem 285: 25103-25108.
- Canalis E, McCarthy T, Centrella M (1988) Growth factors and the regulation of bone remodeling. J Clin Invest 81: 277-281.
- Centrella M, McCarthy TL, Canalis E (1990) Receptors for insulin-like growth factors-I and -II in osteoblast-enriched cultures from fetal rat bone. Endocrinology 126: 39-44.
- Palcy S, Bolivar I, Goltzman D (2000) Role of activator protein 1 transcriptional activity in the regulation of gene expression by transforming growth factor beta1 and bone morphogenetic protein 2 in ROS 17/2.8 osteoblast-like cells. J Bone Miner Res 15: 2352-2361.
- Suzuki T, Nakayama H, Yamaguchi M (1997) Effect of wasabi leafstalk (Wasabia japonica MATSUM.) extract on bone metabolism in mouse calvaria tissue culture. Food Sci Technol Int Tokyo 3: 366-369.
- Suzuki T, Yamaguchi M (1999) Characterization of stimulatory effect of wasabi leafstalk (Wasabia japonica MATSUM.) extract on bone calcification in mouse calvaria tissue culture. Food Sci Technol Res 5: 304-307
- Yohay DA, Zhang J, Thrailkill KM, Arthur JM, Quarles LD (1994) Role of serum in the developmental expression of alkaline phosphatase in MC3T3-E1 osteoblasts. J Cell Physiol 158: 467-475.
- Yohay DA, Zhang J, Thrailkill KM, Arthur JM, Quarles LD (1994) Role of serum in the developmental expression of alkaline phosphatase in MC3T3-E1 osteoblasts. J Cell Physiol 158: 467-475.
- Suzuki M, Yamaguchi M (2003) Stimulatory effect of wasabi leafstalk extract (Wasabia japonica MATSUM.) on bone calcification:
- Interaction with bone anabolic factors in mouse calvaria tissue in vitro.Food Sci Technol Res 9: 87-90
- Suzuki T, Yamaguchi M (1999) Anabolic effect of wasabi leafstalk (Wasabia japonica MATSUM.) extract on bone metabolism in rats. Food Sci Technol Res 5: 61-63
- Yamaguchi M, Ma ZJ, Suzuki T (2003) Anabolic effect of wasabi leafstalk (Wasabia japonica MATSUM.) extract on bone components in the femoral-diaphyseal and -metaphyseal tissues of aged female rats in vitro and in vivo. J Health Sci 49: 123-128
- Yamaguchi M, Uchiyama S, Tsukamoto Y (2002) Stimulatory effect of menaquinone-7 on bone formation in elderly female rat femoral tissues in vitro: prevention of bone deterioration with aging. Int J Mol Med 10: 729-733.
- Suzuki T, Yamaguchi M (2004) Purification of active component in wasabi leafstalk (Wasabia japonica MATSUM) extract in stimulating bone calcification in vitro. J Health Sci 50: 483-490
- Lai YL, Yamaguchi M (2006) Phytocomponent p-hydroxycinnamic acid stimulates bone formation and inhibits bone resorption in rat femoral tissues in vitro. Mol Cell Biochem 292: 45-52.
- Yamaguchi M, Lai YL, Uchiyama S, Nakagawa T (2008)
- Yamaguchi M, Lai YL, Uchiyama S, Nakagawa T (2008) Phytocomponent p-hydroxycinnamic acid stimulates mineralization in osteoblastic MC3T3-E1 cells. Int J Mol Med 22: 287-291.
- Zaidi M, Blair HC, Moonga BS, Abe E, Huang C L-H (2003)
- Zaidi M, Blair HC, Moonga BS, Abe E, Huang CL (2003) Osteoclastogenesis, bone resorption, and osteoclast-based therapeutics. J Bone Miner Res 18: 599-609.
- Tanaka S, Nakamura I, Inoue J, Oda H, Nakamura K (2003) Signal transduction pathways regulating osteoclast differentiation and function. J Bone Miner Metab 21: 123-133.
- Zaidi M, Blair HC, Moonga BS, Abe E, Huang CL (2003) Osteoclastogenesis, bone resorption, and osteoclast-based therapeutics. J Bone Miner Res 18: 599-609.
- Klein-Nulend J, Fall PM, Raisz LG (1990) Comparison of the effects of synthetic human parathyroid hormone (PTH)-(1-34)-related peptide of malignancy and bovine PTH-(1-34) on bone formation and resorption in organ culture. Endocrinology 126: 223-227.
- Takahashi N, Yamana H, Yoshiki S, Roodman GD, Mundy GR, et al. (1988) Osteoclast-like cell formation and its regulation by osteotropic hormones in mouse bone marrow cultures. Endocrinology 122: 1373-1382.
- Lai YL, Yamaguchi M (2007) Phytocomponent p-hydroxycinnamic acid inhibits osteoclast-like cell formation in mouse bone marrow cultures. Int J Mol Med 19: 123-128.
- Nanes MS (2003) Tumor necrosis factor-alpha: molecular and cellular mechanisms in skeletal pathology. Gene 321: 1-15.
- Teti A, Colucci S, Grano M, Argentino L, Zallone AZ (1992) Protein kinase C afftects microfilaments, bone resorption, and [Ca2+] sensing in cultured osteoclasts. Am J Physiol 263:C130-C139
- Franzoso G, Carlson L, Xing L, Poljak L, Shores EW, et al. (1997) Requirement for NF-kappaB in osteoclast and B-cell development. Genes Dev 11: 3482-3496.
- Boyce BF, Xing L, Franzoso G, Siebenlist U (1999) Required and nonessential functions of nuclear factor-kappa B in bone cells. Bone 25: 137-139.
- Iotsova V, Caamaño J, Loy J, Yang Y, Lewin A, et al. (1997) Osteopetrosis in mice lacking NF-kappaB1 and NF-kappaB2. Nat Med 3: 1285-1289.
- Li Y, Li A, Strait K, Zhang H, Nanes MS, et al. (2007) Endogenous TNFalpha lowers maximum peak bone mass and inhibits osteoblastic Smad activation through NF-kappaB. J Bone Miner Res 22: 646-655.
- Eliseev RA, Schwarz EM, Zuscik MJ, O'Keefe RJ, Drissi H, et al. (2006) Smad7 mediates inhibition of Saos2 osteosarcoma cell differentiation by NFkappaB. Exp Cell Res 312: 40-50.
- Yamaguchi M, Weitzmann MN (2009) The bone anabolic carotenoids p-hydroxycinnamic acid and ß-cryptoxanthin antagonize NF-?B activation in MC3T3 preosteoblasts. Mol Med Reports 2:641-644
- Yamaguchi M, Weitzmann MN (2012) submitted
- Vaira S, Alhawagri M, Anwisye I, Kitaura H, Faccio R, et al. (2008) RelA/p65 promotes osteoclast differentiation by blocking a RANKL-induced apoptotic JNK pathway in mice. J Clin Invest 118: 2088-2097.
- Lai YL, Yamaguchi M (2006) Oral administration of phytocomponent p-hydroxycinnamic acid has anabolic effects on bone calcification in femoral tissues of rats in vivo. J Health Sci 52: 308-312.
- Yamaguchi M, Lai YL, Uchiyama S, Nakagawa T (2008) Oral administration of phytocomponent p-hydroxycinnamic acid prevents bone loss in ovariectomized rats. Mol Cell Biochem 311: 31-36.
- Lai YL, Yamaguchi M (2007) Oral administration of phytocomponent p-hydroxycinnamic acid has a preventive effect on bone loss in streptozotocin-induced diabetic rats. Int J Mol Med 19: 803-807.
- Minkin C (1982) Bone acid phosphatase: tartrate-resistant acid phosphatase as a marker of osteoclast function. Calcif Tissue Int 34: 285-290.
- Shires R, Teitelbaum SL, Bergfeld MA, Fallon MD, Slatopolsky E, et al. (1981) The effect of streptozotocin-induced chronic diabetes mellitus on bone and mineral homeostasis in the rat. J Lab Clin Med 97: 231-240.
- Chen JR, Lazarenko OP, Wu X, Kang J, Blackburn ML, et al. (2010) Dietary-induced serum phenolic acids promote bone growth via p38 MAPK/β-catenin canonical Wnt signaling. J Bone Miner Res 25: 2399-2411.
- Nyomba BL, Verhaeghe J, Thomasset M, Lissens W, Bouillon R (1989) Bone mineral homeostasis in spontaneously diabetic BB rats. I. Abnormal vitamin D metabolism and impaired active intestinal calcium absorption. Endocrinology 124: 565-572.
|
| |
| |