A Study on Temperature Changes in Paleocene-Eocene Boundary based on Planktonic Foraminifera in Binak 4 Subsurface Section in Bushehr Province, Southwest of Iran
Received: 31-Jan-2014 / Accepted Date: 01-Mar-2014 / Published Date: 04-Mar-2014 DOI: 10.4172/2157-7617.1000192
Abstract
This study seeks to investigate and identify Paleocene-Eocene boundary in Binak 4 subsurface section in Bushehr province and the effect of temperature changes on the behaviour of planktonic foraminifera. In order to do so, 13 genera and 46 species from Planktonic foraminifera were identified, and two assemblages comprising warm water foraminiferal assemblage and cool water foraminiferal assemblage were segregated. The study of biological events on foraminifera under investigation utilizing universal biozonation led to 5 biozone and 5 subzone segregates. According to the known fossil assemblages and the defined biozones, age in this study was considered late Paleocene (late Selandian) to latest early Eocene (Ypresian). Biostratigraphic evidences (episodic extinction in cool water foraminiferal assemblages and diversity in warm water foraminiferal assemblages) at depths of 8750-8760 (ft.) are indicative of temperature changes and introducer of a global event termed ‘Paleocene-Eocene Thermal Maximum’ (PETM) at this depth. Moreover, In view of the extinction of late Paleocene index species at depths of 8760-8770 (ft. ) and the emergence of early Eocene index species at depths of 8640-8650 (ft.), Paleocene-Eocene boundary is located at depths of 8750-8760 (ft..) in P5 biozone or P5-E1 biozone.
Keywords: Paleocene-Eocene boundary; Selandian; Ypresian
5953Introduction
Paleocen-Eocene boundary is determined by a global event termed ‘Paleocene Eocene Thermal Maximum’ (PETM) [1-9]. This event has triggered one of the biggest and the greatest temperature changes at the global level, the occurrence of which triggered environmental and climate changes directly influencing Paleocene and Eocene assemblages of fauna and flora [10-14].
So far, Paleocene and Eocene deposits have not been studied in this respect in Iran. Paleocene and Eocene deposits have not been continuously deposited throughout Iran. In North of Iran, in the structural zone of Alborz, Paleocene and Eocene deposits are limited to the southern hillsides of Alborz Mountains, while In southwest of Iran, in Zagros sedimentary basin, most of these deposits have been deposited continuously [15].
The purpose of this study in Paleocene-Eocene boundary is determining Paleocene-Eocene boundary based on index species of planktonic foraminifera and investigating the temperature changes based on foraminiferal assemblages in this interval.
Geological setting
Being located in southwest of Iran, Zagros sedimentary basin is one of the structural sedimentary units in Iran (Figure 1) [16]. This basin comprises lands located in the southwest neoTethys suture [15]. Geographically, Zagros basin is divided into three regions, namely Lurestan, Khuzestan and Fars, and geomorphologically, it is divided into two zones, namely imbricate zone and simply folded zone [17]. Zagros Mountains are limited to imbricate zone and simply folded zone introducing young Tetys facies. Paleocene and Eocene deposits have developed in Zagros basin from west regions of Lurestan and Khuzestan to the south of Fars. Furthermore, these deposits cropped out in coastline of Persian Gulf in Bandar Daylam, Bandar Rig, and some parts of Gheshm Island [15]. Binak 4 subsurface section is 20 kilometres away from Bandar Daylam, being located in Binak oil field in Bushehr province (Figure 2). Paleocene Eocene deposits in this subsurface section introduces deposits of Pabdeh formation (Figure 3) [18], being made up of shale along with some chert in the section. Based on the investigation in the section, Pabdeh formation is of 361 meters thickness and Paleocene-Eocene boundary is located in this formation.
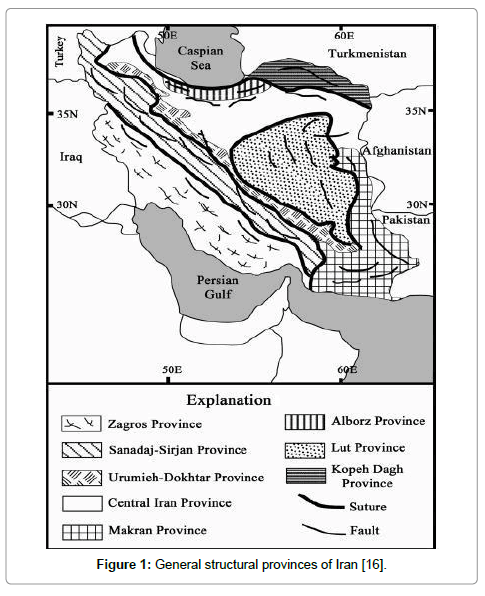
Figure 1: General structural provinces of Iran [16].
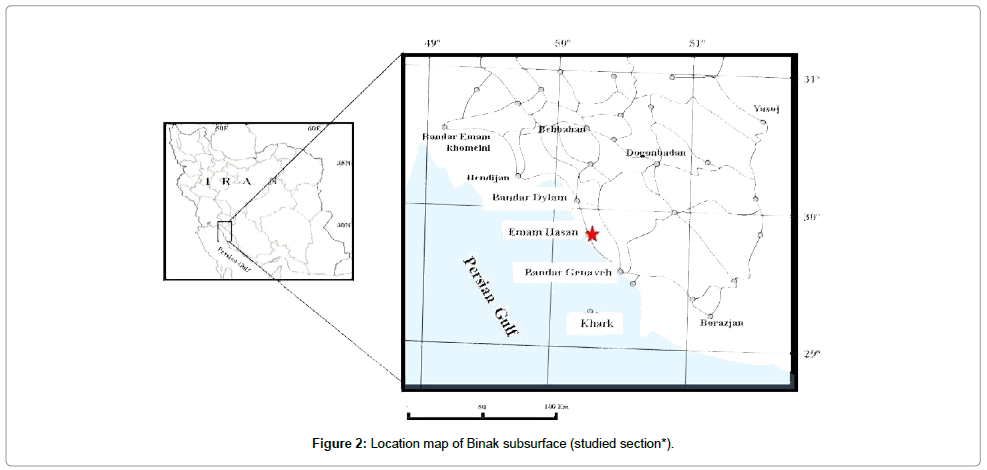
Figure 2: Location map of Binak subsurface (studied section*).
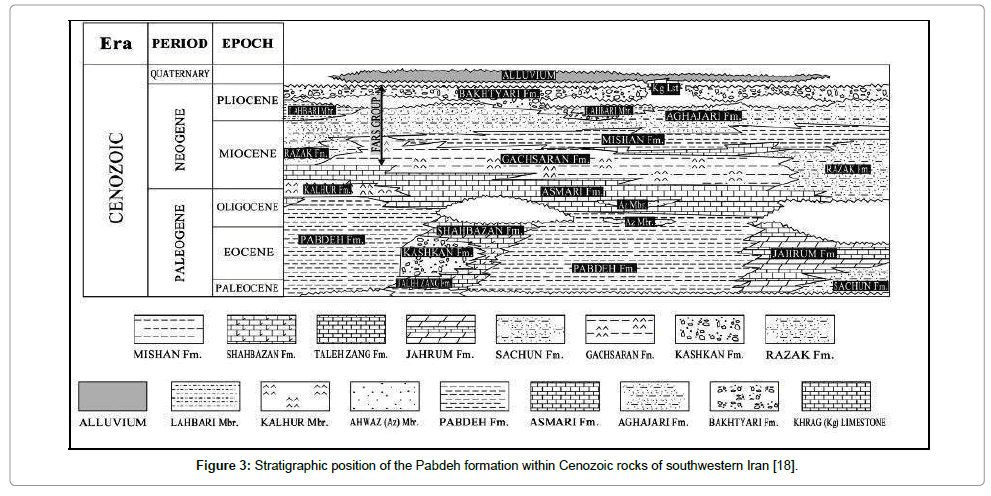
Figure 3: Stratigraphic position of the Pabdeh formation within Cenozoic rocks of southwestern Iran [18].
Material and Methods
In reaching the aim, overall, fine microscopic sections of 120 samples were studied in terms of their contents of planktonic foraminifera. Initially, planktonic foraminifera were systematically identified and named, and their stratigraphic developments were illustrated. Ultimately, they were weighed against the universal biozonation [19,20], and based on this biozonation, age of the sediments were determined. Furthermore, in order to investigate the temperature changes and its effect on planktonic foraminifera of Paleocene and Eocene deposits, the number and the variety of the species found in each of the samples were counted, considering [21] studies on planktonic foraminifera in Paleocene-Eocene boundary, dividing them into two groups of cool water planktonic foraminiferal assemblages and warm water planktonic foraminiferal assemblages; and studying the temperature changes in this boundary taking the proportion of these assemblages into account. Further, types of species pertaining to cool water planktonic foraminiferal assemblages and to warm water planktonic foraminiferal assemblages were segregated. Finally, the proportion of the two assemblages in the sample sequences of Paleocene and Eocene were illustrated.
Results
The systematic study on the foraminifera led to the identification of 13 genera and 46 species of planktonic foraminifera. The study of biological events on planktonic foraminifera in Pabdeh formation on the subsurface section under study utilizing the provided biozonation by Premoli [19] and Wade [20] led to segregation of 5 biozones and 5 subzones. In addition, types of index species in biozones such as Globanomalina psesudomenardii, Morozovella acuta, Acarinina soldadoensis, Morozovella velascoensis, and Morozovella formosa were identified in the section.
P4-Globanomalina pseudomenardii total range zone
This biozone was defined by Boli [22]. The lower boundary and the upper boundary were identified by the appearance and the extinction of Globanomalina pseudomenardii [19]. This biozone is divided into 3 subzones by Berggren [23].
P4a-Acarinina subsphaerica interval range subzone
This subzone ranges from the first appearance of Globanomalina pseudomenardii to the first appearance of Morozovella acuta [19]. In Binak 4 subsurface section, although the first appearance of Globanomalina pseudomenardii was not observed, the upper boundary in this subzone, namely the first appearance of Morozovella acuta is located at depths of 8720-8730 (ft.). It can be assumed that the base of the section with 9.14 meters thickness from Pabdeh formation is located in this subzone. The main planktonic foraminifera in this subzone included in Binak 4 section are Igorina pusilla, Igorina albeari, Morozovella conicutruncata, Morozovella angulata, Acarinina nitida, Acarinina subsphaerica, Morozovella velascoensis, Igorina tadjekestanensis, Morozovella acutispira, and Acarinina mckannai. Accordingly, determined age for this subzone is late Paleocene (late Selandian-early Thanetian).
P4b-Morozovella acuta interval range subzone
This subzone ranges from the first appearance of Morozovella acuta to the first appearance of Acarinina soldadoensis [17]. In Pabdeh formation the lower boundary of this subzone is determined by the first appearance of Morozovella acuta at depths of 8730-8720 (ft.) at the base and the first appearance of Acarinina soldadoensis at depths of 8680- 8690 (ft.) at the top. This subzone is coincident with ‘P4b-Acarinina subsphaerica Partial Range Subzone’ of [20] biozonation. The base of this subzone is not located in the section. Notwithstanding, the upper boundary is marked by the first appearance of Acarinina soldadoensis at depths of 8680-8690 (ft.). This subzone in Pabdeh formation is of 12.9 meters thickness, and the main planktonic foraminifera in this subzone included in Binak 4 section are Acarinina nitida, Acarinina mckannai, Globanomalina pseudomenardii, Morozovella velascoensis, Subbotina triangularis, Subbotina cancellata, and Acarinina esnaensis. Thus, determined age for this subzone is late Paleocene (Thanetian).
P4c-Acarinina soldadoensis concurrent range subzone
This subzone ranges from the first appearance of Acarinina soldadoensis to the last appearance of Globanomalina pseudomenardii [19]. In Pabdeh formation this section is defined by the first appearance of Acarinina soldadoensis at depth of 8680-8690 (ft.) at the base and the last appearance of Globanomalina pseudomenardii at depth of 8660-8670 (ft.) at the top. The current subzone is coincident with ‘P4c- Acarinina soldadoensis/Globanomalina pseudomenardii Concurrent Range Subzone’ of [20] biozonation. This subzone in Pabdeh formation is of 6.9 meters thickness, and the main planktonic foraminifera in this subzone included in Binak 4 section are Igorina albeari, Acarinin nitida, Morozovella velascoensis, Igorina tadjekestanensis, Morozovella acutispira Morozovella acuta, Subbotina triangularis, Acarinina soldadoensis, Subbotina cancellata, and Morozovella aequ. Therefore, determined age for this subzone is late Paleocene (Late Thanetian).
P5-Morozovella velascoensis partial range zone
This biozone is defined by (Boli, 1966) ranges from the last appearance of Globanomalina pseudomenardii to the last appearance of Morozovella velascoensis [19]. In Pabdeh formation, this section defined by the last appearance of Globanomalina pseudomenardii at depths of 8660-8670 (ft.) at the base and the last appearance of Morozovella velascoensis at depths of 8610-8620 (ft.) at the top. This zone is coincident with P5, E1, and E2 zones of [20] biozonation. This zone is determined by the last appearance of Globanomalina pseudomenardii at depths of 8660-8670 (ft.) at the base, and the top of the zone is cannot be identified on account of the index species Acarinina sibaiyaensis being absent. While this zone and E1-Acarinina sibaiyaensis cannot be differentiated and separated, E2-Pseudohastigerina wilcoxensis/ Morozovella velascoensis biozone is recognizable in Binak 4 section and is defined by the first appearance of Pseudohastigerina wilcoxensis in 8640-8650 (ft.) depth at the base, and Morozovella velascoensis at depths of 8610-8620 (ft.) at the top. Also, this zone in Pabdeh formation is of 15.24 meters thickness, and the main planktonic foraminifera in this zone included in Binak 4 section are Globanomalina australiformis, Morozovella Formosa, Morozovella gracilis, Morozovella edgari, Igorina broedmanni, Acarinina wilcoxensis, Pseudohastigerina wilcoxensis, Acarinina soldadoensis, Subbotina velascoensis, Subbotina inaquispira, Acarinina esnaensis, Acarinina intermedia, Morozovella gracilis, Globanomalina chapmani, Morozovella subbotinae, Acarinina decepta, Morozovella velascoensis, Acarinina nitida, Igorina convexa, Acarinina coalingensis, Globanomalina ovalis, and Morozovella aequa. Hence, determined age for this subzone is latest Paleocene-Earliest Eocene (Thanetian to Ypresian).
P6-Morozovella subbotinae partial range zone
This zone is defined by Berggren [23,24] the last appearance of Morozovella velascoensis at the base and the first appearance of Morozovella aragonensis at the top. This zone is divided into two subzones by Berggren [25].
P6a-Morozovella edgari interval subzone
This subzone is defined by the last appearance of Morozovella velascoensis to the first appearance of Morozovella formosa [19]. In Pabdeh formation, this section is defined by the last appearance of Morozovella velascoensis at depths of 8610-8620 (ft.) at the base and the first appearance of Morozovella formosa at depths of 8570-8580 (ft.) at the top. Moreover, this subzone is coincident with ‘E3.Morozovella marginodentata Partial Range Zone’ of [20] biozonation. The main planktonic foraminifera in this subzone included in Binak 4 section are Acarinina nitida, Morozovella aequa, Igorina convexa, Subbotina inaquispira, Acarinina decepta, Acarinina aquiensis, Acarinina intermedia, and Pseudohastigerina wilcoxensis. Accordingly, determined age for this subzone is Earliest Eocene (Ypresian).
P6b-Morozovella lensiformis interval subzone
This subzone is defined by the first appearance of Morozovella formosa at the base and the first appearance of Morozovella aragonensis at the top [19]. In Pabdeh formation, the first appearance of Morozovella formosa at depths of 8570-8580 (ft.) is at the base, and due to the species Morozovella aragonensis’s being absent at the top, the top boundary was determined by the last appearance of Morozovella aequa being located in P6b biozone and the oldest species from genus Globigerinatheka sp. at depths of 8520-8530 (ft.). Therefore, the top of the P6b zone intersecting with the base of P7 biozone is approximately located at depths of 8540-8550 (ft.).
In addition, the current subzone is coincident with ‘E4-Morozovella formosa Lowest Occurrence Zone’ of [20] biozonation, and in Pabdeh formation it is of 13 meters thickness. The main planktonic foraminifera in this subzone included in Binak section are Morozovella aequa, Acarinina collactea, Pseudohastigerina wilcoxensis, Acarinina aquiensis, Acarinina peseudotopilensis, Morozovella formosa, Acarinina intermedia, and Chilogumbelina sp. Accordingly, determined age for this subzone is Early Eocene (Ypresian).
P7-Morozovella Formosa concurrent rang zone
This biozone is defined by Berggren [25] the first appearance of Morozovella aragonensis at the base and the last appearance of Morozovella formosa at the top [19]. In Pabdeh formation, due to the species Morozovella aragonensis’s being absent at the base, the base boundary was determined by the last appearance of Morozovella aequa being located in P6b biozone and the oldest species from genus Globigerinatheka sp. at depths of 8520-8530 (ft.). Therefore, the top of the P6b zone intersecting with the base of P7 biozone is approximately located at depths of 8540-8550 (ft.), and the top of this biozone was determined by the last appearance of Morozovella formosa at depths of 8505-8510 (ft.).
Moreover, the current zone is coincident with ‘E5-Morozovella aragonensis/Morozovella subbotinae Concurrent Range Zone’ of [20] biozonation, and in Pabdeh formation it is of 12 meters thickness. The main planktonic foraminifera in this subzone included in Binak 4 section are Acarinina collactea, Acarinina peseudotopilensis, Morozovella formosa, Chilogumbelina sp., Acarinina primitiva, Globigerinatheka sp., Tenuitella reissi, Acarinina interposita, and Acarinina esnaensis. Therefore, determined age for this subzone is middle Early Eocene (Ypresian).
P8-Morozovella aragonensis interval range zone
This zone is defined by Boli [22] the last appearance of Morozovella formosa at the base and the first appearance of Planorotalites palmerae at the top [19]. In Pabdeh formation the base of this zone is defined by the last appearance of Morozovella formosa at depths of 8505-8510 (ft.), and the top of this zone is not identifiable due to the pertinent samples’ no being at hand. This zone is coincident with ‘E6-Acarinina pentacamerata Partial Range Zone’ of [20] biozonation, and in Pabdeh formation it is of 28 meters thickness. The main planktonic foraminifera in this subzone included in Binak 4 section are Acarinina bullbrooki, Acarinina pentacamerata, Truncorotalides rohri, Acarinina interposita, and Chilogumbelina sp. Thus, determined age for this subzone is late Early Eocene (Ypresian).
This zone is of [20] biozonation. It is of 28 meters thickness, the first 7 meters of which is sampled and the last 27 meters of which is not. The accompanying foraminifera in this zone included in Binak 4 section are Acarinina bullbrooki, Acarinina pentacamerata, Truncorotalides rohri, Acarinina interposita, and Chilogumbelina sp. accordingly, age for this biozone is Early Eocene (Figure 4).

Figure 4: Lithostratigraphy and distribution of planktonic foraminifera in Binak 4 section.
Discussion
Paleocene Eocene boundary has been studied over the past decade by [1,4,5,10-14,21,22,26-32]. These studies indicate that one of the foremost climate changes, namely the global warming event, occurred in Paleocene-Eocene boundary. This event occurred approximately 56 million years ago, and was accompanied by many ecological and environmental changes and changes in fauna and flora assemblages. Accumulation of greenhouse gases in the atmosphere, increased humidity, and extinction of and significant decrease in planktonic foraminiferal assemblages and benthic foraminiferal assemblages are among these changes in Paleocene-Eocene boundary [1,4-6,8].
In planktonic foraminifera, the last appearance of Igorina albeari which coincides with the first appearance of Acarinina sibaiyaensis and Acarinina Africana, and the simultaneity in the extinction of benthic foraminiferal assemblages along with climate changes were selected as markers for Paleocene-Eocene boundary [8,21].
Many scholars have carried out research on this boundary and the effects of climate changes on planktonic and benthic foraminiferal assemblages. Lu [8] studied on the number and the variety of planktonic foraminifera in the Paleocene-Eocene boundary in Spain. They divided foraminiferal assemblages into three foraminiferal assemblages based on depth of location, i.e. surface foraminiferal assemblages comprising genera Acarinina, Morozovella, and Igorina; subsurface foraminiferal assemblages comprising genera Subbotinae, Chilogumbelina and Pesudohastigerin; and deep foraminiferal assemblages comprising genus Globanomalina. They studied the abundance and variety in these assemblages in sequences leading to the Paleocene-Eocene boundary extremes. According to Lu [8], abrupt warming of the waters resulted in significant decrease and episodic extinction in the abundance of subsurface and deep assemblages lived in cool water and susceptible to this abrupt warming. By contrast, surface foraminiferal assemblages being environmentally compatible became diverse and abundant. In this study two genera, namely Acarinina and Morozovella, were of dominant genera in Paleocene-Eocene boundary.
Pardo [21] also studied the scattering pattern of planktonic foraminifera in Paleocene Eocene sequences in Tethys basin, and came to conclusions respecting Paleocene-Eocene boundary. They divided foraminiferal assemblages into two groups of warm water foraminiferal assemblages comprising genera Acarinina, Morozovella, and Igorina; and cool water assemblages comprising genera Subbotina, Chilogumbelina, Pesudohastigeria, and Globanomalina. They believed that owing to climate changes and global warming, cool water foraminiferal assemblages, thus becoming environmentally incompatible, became extinct. By contrast, abundance in warm water foraminiferal assemblages triggered their immigration from lower latitudes to higher ones (Figure 4).
Pardo [21] also studied the scattering pattern of planktonic foraminifera in Paleocene Eocene sequences in Tetys basin, and came to conclusions respecting Paleocene-Eocene boundary. They divided foraminiferal assemblages into two groups of warm water foraminiferal assemblages comprising genera Acarinina, Morozovella and Igorina; and cool water assemblages comprising genera Subbotina, Chilogumbelina, Pesudohastigeria and Globanomalina.
In the studies of Pardo [21] genus Acarinina had more compatibility with this temperature changes compared to other genera from warm water foraminiferal assemblages. By contrast, genus Morozovella was less compatible with this global warming, thus the number and the variety of its species decreased.
In the selected section under study, taking the provided biozonation and its being coincident with the universal biozonation [19,20] into account, Paleocene-Eocene boundary is located in deposits of Pabdeh formation.
Based on the evidences available at the section under investigation, the last appearance of Igorina albeari at depths of 8760-8770 (ft.) in P5 biozone as a marker for the late Paleocene prior to Paleocene-Eocene boundary and the first appearance of the genera Acarinina intermedia, Pesudohastigerina wilcoxensis, and Morozovella gracilis in P5 or E2 at depths of 8640-8650 (ft.) were identified as early Eocene first species. Therefore, taking these fossil evidences into account, this boundary is located at depths of 8750-8760 (ft.) in P5 biozone or P5-E1 biozone. Moreover, the effect of global event of PETM at this depth and its temperature changes effects on foraminiferal assemblages corroborate the Paleocene-Eocene boundary to be located at depths of 8750-8760 (ft.) in P5 biozone or P5-E1 biozone.
In the analysis of the development and variety diagram of planktonic foraminifera in Pabdeh formation, taking Pardo [21] studies into account, two foraminiferal assemblages of warm water and cool water were identified. These two assemblages lie in the same group with those defined by Lu [8] based on depth of foraminifera locations. Warm water foraminiferal assemblages are the same surface foraminiferal assemblages, and cool water foraminiferal assemblages are the same subsurface and deep foraminiferal assemblages. In the initial sequences of Pabdeh formation, both assemblages were observed with abundance and variety in species.
Types of species pertaining to Acarinina and Morozovella from warm water foraminiferal assemblages and those pertaining to Subbotina and Globanomalina from cool water assemblages with high abundance reside in P4 biozone.
In the section under study, there was a dramatic decrease in the variety of planktonic foraminifera at depths of 8750-8760 (ft.), and significant changes occurred at this depth in warm water and cool water foraminiferal assemblages. From cool water foraminiferal assemblages, species such as Subbotina triangularis, Subbotina velascoensis, Subbotina inaquispira, Globanomalina ovalis and Globanomalina chapmanii became extinct, and from warm water foraminiferal assemblages only species of Acarinina nitida, Acarinina coalingensis, Acarinina decepta, (Figure 5) [33,34] and Acarinina esnaensis were observed at this depth. Species of the genus Morozovella from warm water foraminiferal assemblages being abundant up to this depth became extinct as Morozovella velascoensis, Morozovella acuta and Morozovella aequa did at this depth. Only Morozovella marginodentata, Morozovella Formosa and Morozovella aequa were observed in upper Eocene sequences.
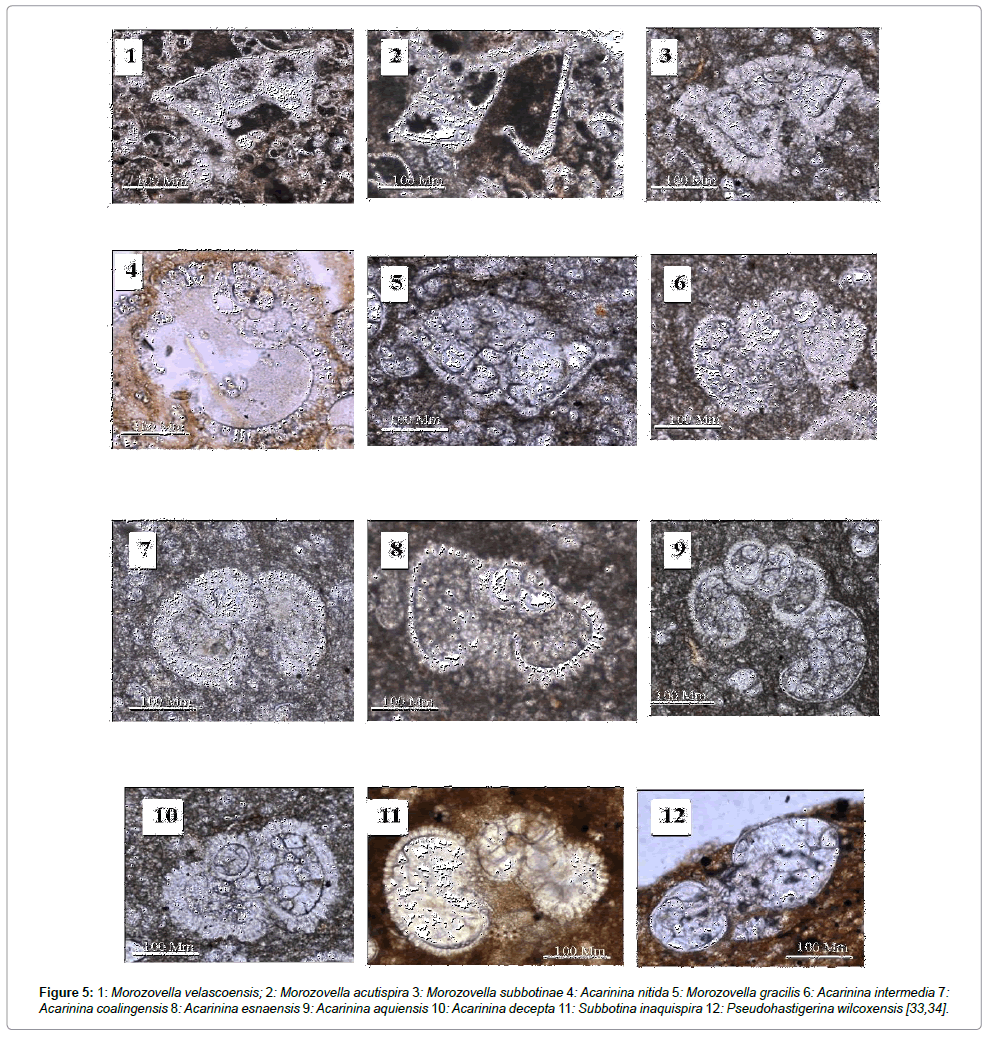
Figure 5: 1: Morozovella velascoensis; 2: Morozovella acutispira 3: Morozovella subbotinae 4: Acarinina nitida 5: Morozovella gracilis 6: Acarinina intermedia 7: Acarinina coalingensis 8: Acarinina esnaensis 9: Acarinina aquiensis 10: Acarinina decepta 11: Subbotina inaquispira 12: Pseudohastigerina wilcoxensis [33,34].
Passing from this depth through, abundance and variety gradually increases in cool water foraminiferal assemblages in P6a, P6b, and P7 biozones in Eocene sequences deposits.
Evidences also demonstrate that temperature changes and thermal fluctuations at depths of 8750-8760 (ft.) in late Paleocene and early Eocene triggered this different behaviour in planktonic foraminifera. Cool water foraminiferal assemblages being incompatible with water are being warmed at this depth became episodically extinct. By contrast, warm water foraminiferal assemblages having favourable conditions for living and flourishing increased, being the only genera observed at this depth. Moreover, temperature changes at this depth correspond with temperature changes occurred by virtue of the global event PETM in Paleocene-Eocene boundary. Therefore, depths of 8750-8760 (ft.), being located in P5 biozone and being encompassed by late Paleocene index species in older sequences and by early Eocene index species in younger sequences, is introduced as Paleocene-Eocene boundary.
It is to mention that in the studies of Pardo [21] and Lu [8] on the transitional boundary, in addition to the types of species of genus Acarinina, the types of species of genera Morozovella and Igorina were reported to be from warm water foraminiferal assemblages, while in Paleocene-Eocene boundary in the section under investigation, we observed a decrease in the abundance and the variety in warm water foraminiferal assemblages. The difference in the abundance of warm water foraminiferal assemblages is presumed to be due to Iran’s being located in the higher latitudes.
Conclusion
For the initial part of Pabdeh formation in Binak 4 subsurface section, 13 genera and 46 species from planktonic foraminifera were identified. In the late Paleocene-early Eocene boundary based on Premoli [19] and Wade [20] biozonation, 5 biozones and 5 subzones were identified. Identified fossil assemblages, the area under investigation is late Paleocene to latest early Eocene. Out of 104 meters thickness of the lower part of Pabdeh formation, 31 meters is Paleocene deposits and 73 meters is Eocene ones.
Biostratigraphic evidences in the section under investigation indicate that the global event PETM is located in P5 zone at depths of 8750-8760 (ft.). This global event introducing Paleocene-Eocene boundary has influenced planktonic foraminiferal assemblages of this formation. The behaviour of planktonic foraminiferal assemblages was different towards PETM. Warm and cool planktonic foraminiferal assemblages were identified. At 8650-8660 ft. only warm planktonic foraminiferal assemblages were found. Post these depth only cool planktonic foraminiferal assemblages were abundant. Considering these evidences, Paleocene-Eocene boundary in Pabdeh formation sediments in Binak 4 subsurface section being under investigation was determined to be at depths of 8650-8660 (ft.). Genus Morozovella is temperature sensitive and majority of its genera got extinct in Paleocene-Eocene boundary.
In view of the fact that the last appearance of the genus Igorina albeari at depths of 8760-8770 (ft.) in P5 biozone was identified as a marker for late Paleocene before Paleocene-Eocene boundary, and the first appearance of the genera Acarinina intermedia, Pesudohastigerina wilcoxensis and Morozovella gracilis in P5 or E2 at depths of 8640-8650 (ft.) was identified as early Eocene first species, taking these fossil evidences into account, this boundary resides at depths of 8750-8760 (ft.) in P5 biozone or P5-E1 biozone. Furthermore, the effects of the global event PETM at this depths being universally concurred to have occurred in Paleocene-Eocene boundary combined with the effects of temperature changes on foraminiferal assemblages substantiate the Paleocene-Eocene boundary’s being located at depths of 8750-8760 (ft.) in P5 biozone or P5-E1 biozone.
References
- Stassen P, Thomas E, Speijer RP (2012) Restructuring outer nerictic foraminiferal assemblages in the aftermath of the Paleocene-Eocene thermal maximum. J Micropal 37: 89-93.
- Zachos JC1, Röhl U, Schellenberg SA, Sluijs A, Hodell DA, et al. (2005) Rapid acidification of the ocean during the Paleocene-Eocene thermal maximum. Science 308: 1611-1615.
- Zachos JC, Schouten S, Bohaty S, Quattlebaum T, Sluijs A, et al. ( 2006) Extreme warming of mid-latitude coastal ocean during the Paleocene–Eocene Thermal Maximum: inferences from TEX86 and isotope data. Geol 34: 737-740.
- Zachos JC, Dickens GR, Zeebe RE (2008) An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 451: 279-283.
- Guasti E, Speijer PR (2008) Acarininamulticameratan.sp. (Foraminifera): a new marker for the Paleocene Eocene thermal maximum. J Micropal 27: 5-12.
- Kelly DC, Bralower TJ, Zachos JC, Premoli SI, Thomas E (1996) Rapid diversification of planktonic foraminifera in the tropical Pacific ODP Site 865 during the Late Paleocene Thermal Maximum. Geol 24: 423-426.
- Kelly DC, Bralower TJ, Zachos JC ( 1998) Evolutionary consequences of the latest Paleocene thermal maximum for tropical planktonic foraminifera. PalaeogeoPalaeoclimatPalaeoecol 141: 139-161.
- Lu G, Keller G, Pardo A (1998) Stability and change in Tethyanplanktic foraminifera across the Paleocene-Eocene transition. Marine Micropal 35: 203-233.
- Arenillas I, Molina E (2000) Reconstrucciónpaleoambiental con foraminÃferosplanctónicos y cronoestratigrafÃadeltránsitoPaleoceno–Eoceno de Zumaya (Guipúzcoa). Revista Española de Micro 32: 283-300.
- Alegret L, Ortiz S (2007) Global extinction event in benthic foraminifera across the Palocene/Eocene boundary at the Dababiyastratotype section.Micro 52: 433-447.
- Alegret L, Ortiz S, Arenillas I, Molin E (2010) What happens when the ocean is overheated? The foraminiferal response across the Paleocene–Eocene Thermal Maximum at the Alamedilla section (Spain). GeolSoci of America B 122: 1616-1624.
- Westerhold T, Rohl U, Raffi I, Fornaciari E, Monechi S, et al. (2008) New chronology for the late Paleocene thermal maximum and its environmental implications. Palaeogeo, Paleoclimat, Palaeoecol 257: 377-403.
- Ouda K (2003) The Paleocene/Eocene boundary in Egypt: An overwiew .Micopaleo 99: 15-40.
- Cohen AS, Coe AL, Kemp DB (2007) The late Palaeocene –Early Eocene and Toarcian (Early Jurassic) carbon isotope excursions: a comparison of their time scales, associated environmental changes, causes and consequences. J GeolSoc 164: 1092-1108.
- Motiei H (1994) Geology of Iran. Stratigraphy of Zagros, Geological survey of Iran, Iran.
- Heydari E, Hassanzadeh J , Wade WJ , Ghazi AM (2003) Permian–Triassic boundary interval in the Abadeh section of Iran with implications for mass extinction, Part 1–Sedimentology. PaleogeoPaleoclimatPaleoecol 193:405-423.
- Alavi M (2004) Regional stratigraphy of zagros fold-thrust belt of Iran and its proforeland evolution. American J Sci 304: 1-20.
- Motiei H (2001) Simplified table of rock units in southwest Iran. Tehran, Keyhan Exploration and Production services, Iran.
- Premoli SI, Rettori R, Verga D (2003) Practical Manual of Paleocene and Eocene Plaktonic Foraminifera. International School on Planktonic Foraminifera Perugia,University of Perugia, Italy.
- Wade BS, Pearson PN, Berggren WA, Palike H (2011) Review and revision of Cenozoic tropical plaktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale. Earth Sci Rev 104: 111-142.
- Pardo A, Keller G, Oberhansli H (1999) Paleoecologic and Paleoceanographic evolution of the Tethyan Realm during the Paleocene-Eocene Transition. J Foraminiferal Res 29:37-57.
- Bolli HM (1966) Zonation of Cretaceous to Pliocene Marin Sediments based on Planktonic Foraminifera.GeologischesInstitut der Eidg. TechnischenHochschule und der Universität Zürich, Switzerland.
- Berggren WA, Kent DV, Swisher CC, Aubry MP (1995) A revised Cenozoic Geochronology and Chronostratigraphy. In: Geocronology time scale and Global stratighraphic correlation.Soc Econ Paleontol Mineral Spec Publo 54: 129-212.
- Berggren WA, Pearson P (2005) A Revised tropical to subtropical Paleogene planktonic foraminiferal zonation. J Foraminiferal Res 35: 279-298.
- Berggren WA, Aubry MP, Van Fossen MP, Kent DV, Norris RD, et al. (2000) Tntegrated Palaeocene calcareous plankton magnetobiochronology and stable isotope stratighraphy: DSDP Site 384(NW Atlantic Ocean). Palaeogeo Palaeoclimat Palaeoecol 159: 1-51.
- Berggren WA, Miller KG (1988) Paleogene tropical planktonic foraminiferal biostratigraphy and magnetobiochronology. Micropal 34: 362-380.
- Canudo JI, Molina E (1992) Planktic foraminiferal faunal turnover and biochronostratigraphy of the Paleocene–Eocene boundary at Zumaya, northern Spain. Revista de la Sociedad Geológica de España 5: 145-157.
- Lu G, Keller G (1995) Planktic foraminiferal faunal turnovers in the subtropical Pacific during the late Paleocene to early Eocene. J Foraminiferal Res 25: 97-116.
- Lu G, Keller G (1995) Ecological stasis and saltation: species richness change in planktic foraminifera during the late Paleocene to early Eocene, DSDP site 577. Palaeogeogr Palaeclimatol Palaeoecol 117: 211-227.
- Lu G, Keller G (1996) Seprating ecological assemblages using stable isotope signals: late Paleocene to early Eocene planktic foraminifera, DSDP Site 577. J Foraminiferal Res 26: 103-112.
- Thomas DJ, Bralower TJ, Zachos JC (1999) New evidence for subtropical warming during the late Paleocene thermal maximum: Stable isotopes from Deep Sea Drilling Project Site 527, Walvis Ridge. Paleoceano 14: 561-570.
- RodrÃguez-Tovar FJ, Uchman A, Alegret L, Molina E (2011) Impact of the Paleocene–Eocene Thermal Maximum on the macrobenthic community: Ichnological record from the Zumaia section, northern Spain. Marine Geol 282: 178-187.
- Bolli HM (1957) Planktonic foraminifera from the Eocene Navet and San Ferando formation of Trinidad B.W.L. US Nat Mus Bull 215: 155-172.
- Loeblich AR, Tappan H (1988) Foraminiferal Genera and their Classification. Van Nostrand Reinhold Company Inc, USA.
Citation: Moradian F, Baghbani D, Daneshian J, Allameh M (2014) A Study on Temperature Changes in Paleocene-Eocene Boundary based on Planktonic Foraminifera in Binak 4 Subsurface Section in Bushehr Province, Southwest of Iran. J Earth Sci Clim Change 5: 192. DOI: 10.4172/2157-7617.1000192
Copyright: ©2014 Moradian F, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Share This Article
Recommended Journals
Open Access Journals
Article Tools
Article Usage
- Total views: 15843
- [From(publication date): 4-2014 - Nov 21, 2024]
- Breakdown by view type
- HTML page views: 11224
- PDF downloads: 4619