Use of CLIMEX, Land use and Topography to Refine Areas Suitable for Date Palm Cultivation in Spain under Climate Change Scenarios
Received: 13-Jun-2013 / Accepted Date: 10-Jul-2013 / Published Date: 15-Jul-2013 DOI: 10.4172/2157-7617.1000145
Abstract
In this study, CLIMEX modeling software was used to develop a model of the potential distribution of P. dactylifera under current and various future climate scenarios for Spain. CLIMEX parameters were adjusted depending on satisfactory agreement between the potential and known distribution of P. dactylifera in northern African countries, Iraq, Saudi Arabia, Oman and Iran. The potential date palm distribution was modeled under current and future climate scenarios using one emission scenario (A2) with two different Global Climate Models (GCMs): CSIRO-Mk3.0 (CS) and MIROC-H (MR). The CLIMEX outputs were then refined by land use types and areas less than 10̊ slope, since sloping areas impose problems in hydraulic conductivity and root development. The refined results indicated that large areas in Spain are projected to become climatically more suitable for date palm growth by 2100. However, the results from the CS and MR GCMs show some disagreements. The refined MR GCM projected that approximately 22.86 million hectares in Spain may become suitable for date palm growth, while the CS GCM showed approximately 18.72 million hectares by 2100. The refined results showed that only about 65% of CLIMEX results are suitable for date palm cultivations while the rest of the areas are unsuitable due to the unsuitability of land uses and slope. Our results indicated that cold and wet stresses will play a significant role in date palm distribution in some central and northern regions of Spain by 2100.
Keywords: Climate change; CLIMEX; Phoenix dactylifera L; Date palms
10133Introduction
Global climate change resulting from changes in air and sea surface temperatures [1]; precipitation patterns, ocean level alterations [2] and ocean salinity [3,4] can affect agricultural productivity [5-7]. A review of literature reveals there have been many studies on the effect of climate change on ecological plant processes, such as plant growth [8], crop yield [9] and plant community interactions [3,10-14]. While McDermott [15] documented a greater than 55% crop failure in agricultural regions in India as a consequence of climate change, a comparable study reports that Sudan experienced a 50% crop failure in 2009 as a consequence of small changes in the quantity and pattern of precipitation, resulting from climate change [16]. Further, the potential effect on maize production of climate change has been reported [17] with indications of a 10% reduction by 2055, resulting in a loss of $2 billion per year in Africa and Latin America [17]. Shabani et al. [18] have reported on the effect of altered climate on global date palm production, in terms of long-term broad-scale shifts in the areas conducive to date palm cultivation. The total annual income from date palms in the Middle Eastern countries decreased from 1990 to 2000, due to water shortage and plant diseases resulting from climate change [19]; while an earlier example indicates that climate change caused a $438 million loss in wheat, a $116 million loss in grapes and a $67 million loss in sugar production in Australia and North America [14]. Substantial challenges for development and food security are posed by such losses in production and necessitate modeling the effects of climate change on agricultural production, especially crops important economically, to study the probable impact on agricultural output and distribution.
Date palm, which includes approximately 400 different species within the Arecaceae family, is a crop of economic importance in Middle Eastern countries, including Saudi Arabia, Egypt, Iran, Iraq, as well as some European countries, such as Turkey and Spain [20-24]. Date palm, being suited to a Mediterranean climate with salty and alkaline soils, has been grown in Spain for over 150 years and has become one of the main income sources for local farmers and the Spanish government [25]. Elche, located on the east coast of Spain, is one of Europe’s largest date palm plantations [26,27]. A statistical report indicates that date palm production in Spain reached 3,732 metric tons in 2003 [28]. Climatic parameters, plant diseases resulting from climate change and water shortage are factors that will impact on future date palm yields. Thus identifying how different regions of Spain may be affected by climate change is vital in terms of the future of date palm production in Spain.
Global climate models (GCMs) are important analytical tools in such disciplines as biogeography, evolution, ecology, conservation and invasive species management [29] and extensive use has been made of GCMs in biogeography, conservation biology and environmental management studies since 1996 [3,8,18,30-34]. The major function of climatic models is to illustrate plant sensitivity to climate change, through ascertaining relationships between plant and atmospheric sciences [3]. Such models use the current climate range to identify climate tolerance limits of particular species. Then, by modeling climate change scenarios, the alteration of the areas suitable for growth of vulnerable species can be projected into the future [35]. Such projections are valuable for the development of agricultural organizations’ long-term management strategies for sustainable production of economically important crops such as date palm (Phoenix dactylifera L.) under predicted future climate conditions [18].
There have been many models used to address the climate change factors, including bioclimatic models, ecological niche models (ENMs) [36], species distribution models (SDMs) [36-38] and CLIMEX [39]. CLIMEX is a computer-based software, designed for matching climates in ecology and has been widely used in estimating potential geographical distribution of different species in the future [11,40-44]. With CLIMEX, users can identify areas where selected species could establish and maintain or develop, based on predicted climate alterations. Through comparisons with other correlative modeling methods [45], CLIMEX has been evaluated as the most successful climate modeling software, for describing species’ responses to climate, due to its geographic range and powerful phonological observations [18,40,46,47].
Criticism of CLIMEX indicates that it does not include biotic interactions and dispersal in the modeling process [10,13]. Thus CLIMEX modeling output is based on limited climatic variables, such as overall maximum and minimum monthly temperatures (Tmax and Tmin), overall monthly precipitation levels (Ptotal) and the relative humidity taken at 09:00 h (RH09:00) and 15:00 h (RH15:00) [46,48]. This means that CLIMEX outputs are only based on climate predictions and the projected areas may still be unsuitable due to variables such as soil types, high slopes and urban areas. Thus, refining date palm suitability regions modeled with CLIMEX by investigating slopes and land use would result in a more accurate identification of the areas available for future date palm cultivation. Such refining can be undertaken using geographic information systems (GIS), remote sensing software (RS), species distribution models (SDMs) and ecological niche models (ENMs) [18,30,49-53]. In this study, it has been proposed that the distribution of P. dactylifera may effectively change as a result of climate alteration. With this possible outcome in mind, this research made use of the CLIMEX software package to determine the potential distribution of P. dactylifera under current and future climate and the location of the suitable slopes and land uses to assess the sensitivity of its distribution to climate change, and to evaluate the concurrent implications for cultivation of date palms for 2030, 2050, 2070 and 2100 in Spain.
Methodology
Distribution of date palms (P. dactylifera)
Landsat images, with 30 m image resolution [54], the Global Biodiversity Information Facility (GBIF) [55], Missouri Botanical Gardens’ database [56] and other date palm literatures in CAB Abstracts databases [57] were used to collect data on P. dactylifera distribution and this information was supplemented by other date palm literature [22,24,58-70]. The GBIF database for Northern African countries, Saudi Arabia, Oman, Iraq and Iran had 314 records for P. dactylifera, but 64 records did not have geographical coordinates and were therefore removed, leaving 250 records. Some of these records were duplicates and were also deleted. In the end, 147 records from the GBIF database, 33 records from Missouri Botanical Gardens’ database and 43 records obtained from literature research were used to validate parameters in CLIMEX.
CLIMEX Software, climate data, climate models and climate scenarios
As one of the most comprehensive inferential modeling software packages, CLIMEX allows users to project the climatic impact on potential distribution of different species, based primarily on their current distribution [71]. The program makes use of known associations among climate, species distribution and their biological responses to project their future distributions [72]. Experimental observations of species’ growth response to temperature and soil moisture can be taken into account in CLIMEX [8,30,73,74]. The Ecoclimatic Index (EI) in CLIMEX is an average yearly index of climatic suitability based on regular weekly calculations of growth and stress indices. EI values range from 0 to 100, and a species can be established in an area if EI>0. EI values ranging from 0 to 10 indicate marginal potential habitats, while EI values >20 are highly favorable predictors [40]. Thus, an area with an EI>20 represents a potential area of high suitability from a climatic perspective, for introducing a particular species.
A variety of climatic parameter averages recorded between 1950 and 2000 are include in the CliMond 10’ gridded climate records and these were used to model the current distribution of date palm [46]. The meteorological database’s climatic parameters include overall maximum and minimum monthly temperature (Tmax and Tmin), overall monthly precipitation (Ptotal) and relative humidity as recorded at 09:00 h (RH09:00) and 15:00 h (RH15:00). These parameters were also used to predict climates for the mid- to late 21st century. The potential distribution of date palms under future climate was modeled using two Global Climate Models (GCMs), namely, CSIRO-Mk3.0 and MIROC-H (Center for Climate Research, Japan), with the A2 SRES scenario. These two GCMs were part of the CliMond dataset and were selected from 23 GCMs based on criteria such as the availability of required variables (including temperature, precipitation, sea level pressure and humidity for CLIMEX), small horizontal grid spacing in both GCMs and better representation of observed climate at local scales, compared with other GCMs [2,75,76].
The A2 SRES scenario was chosen as a possible climate scenario principally because of its inclusion of demographic and financial factors, as well as incorporating technological forces driving GHG emissions. The A2 scenario assumptions take into account factors of independent and self-reliant nations, such as population increases and regional economic development [77]. Future assumptions inherent in A2 SRES include high population growth, coupled with slow economic growth and extensive technological change. The A2 scenario assumes less extreme range of GHG emissions than scenarios such as A1F1, A1B, B2, A1T and B1 [77,78].
Model framing
The global distribution of date palm and the suitable areas for date palm cultivation at a global scale have been recently reported by Shabani et al. [18]. The reported CLIMEX parameters were used for Northern African countries, Saudi Arabia, Oman, Iraq, and Iran as the validation areas. Here, temperature and moisture response parameter values were obtained from the date palm physiology and growth model literature [16,22,35,58,67,68,79,80] and transformed into CLIMEX-compatible Temperature and Moisture Index parameters and cold, heat, dry and wet stress threshold values. The final values used for climatic parameters and growth indices in CLIMEX were 0.9 for the wet stress threshold (SMWS), 0.022 week–1 for the accumulation rate of wet stress (HWS), 4°C for the cold stress temperature threshold (TTCS), –0.01 week–1 for the frost stress accumulation rate (THCS), 46°C for the heat stress parameter (TTHS), 0.9 week–1 for the heat stress accumulation rate (THHS), 14°C for the limiting low temperature (DV0), 20°C for the lower temperature (DV1), 39°C for upper optimal temperatures (DV2), 46°C for the limiting high temperature (DV3), 0.007 for the limiting low moisture threshold (SM0), 0.013 for the optimal soil moisture (SM1), 0.81 for the upper optimal soil moisture thresholds (SM2) and 0.9 for the limiting high soil moisture (SM3) [24,25,55,58,62,65,70]. The data sets were output from CLIMEX and imported into ArcGIS (GIS) software for further processing and mapping.
Refining the CLIMEX outputs by suitable land uses and slopes
All vector data representing unsuitable land uses in Spain were taken from European Environment Agency Datasets [81]. The digital elevation model (DEM) of Spain on a regular 30 m grid from ASTER [82] (NASA, 2012) database were used to generate a slope surface, which was then reclassified into the two slope categories, mainly because overlaying the satellite images onto the date palm sites showed that 90% of the locations were on slopes less than 10 degree slopes (suitable 10 degree). Slope classes in raster format were converted to polygon shape files and queries were undertaken using attribute and location of unsuitable land uses including construction sites, coniferous forest, mineral extraction sites, rice fields, salt marshes, road and rail networks and associated land, green urban areas and bare rocks to extract the areas having suitable slopes and suitable land uses. The CLIMEX outputs for current time were overlaid on the location of suitable slopes and suitable land uses for the whole country and all those locations that satisfied the condition of EI>20 were selected.
Results
Model validation under current climate
The distribution of P. dactylifera taken from different databases and an appropriate match between the Ecoclimatic Index (EI) from the CLIMEX model for Morocco, Algeria, Libya, Egypt, Saudi Arabia, Oman, Iraq and Iran is illustrated in Figure 1. The modeled results showed that southern Iran, Oman, Saudi Arabia, northern Egypt, northern Libya, Algeria and western Morocco have suitable climatic condition for P. dactylifera. While large parts of Libya and Algeria are modeled to have suitable climatic conditions for P. dactylifera in its current known distribution, inadequate distribution data were available from these regions. This could be due to a shortage of reporting from these areas as a result of lack of human distribution, or biotic factors such as competition or lack of dispersal opportunities could prevent this species from occurring in these areas [31,83-85]. Nearly 89% of the occurrence records fell within the suitable categories, confirming that the values selected for the various parameters in CLIMEX were optimum (Figure 1).
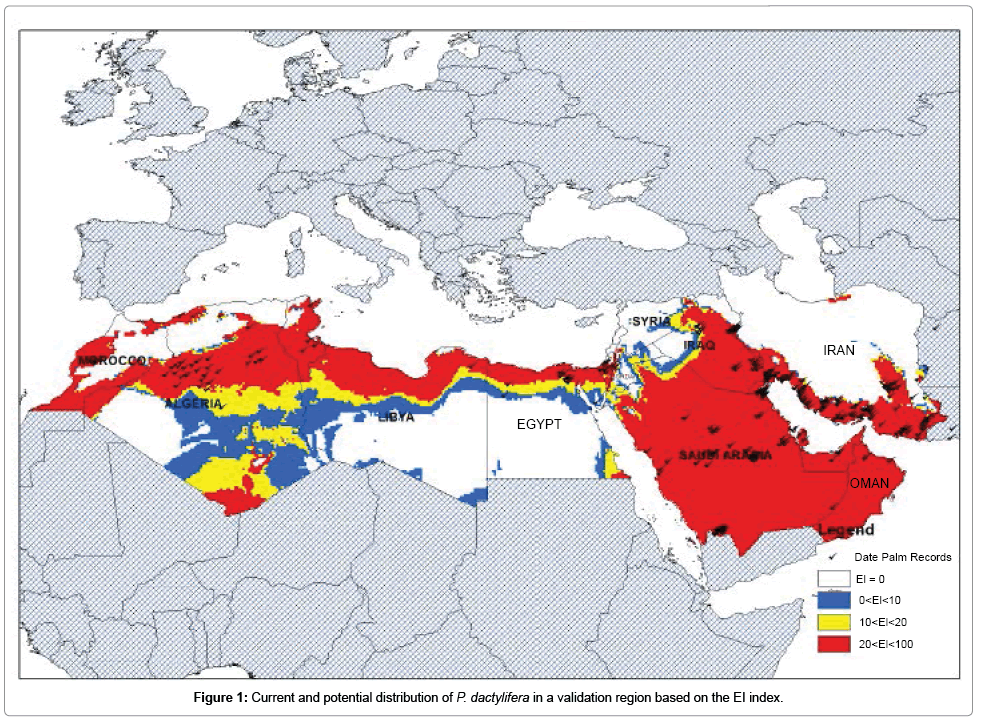
Figure 1: Current and potential distribution of P. dactylifera in a validation region based on the EI index.
Future climate
Figures 2 and 3 are illustrations of the Ecoclimatic Index for P. dactylifera, using the two global climate change models (CS and MR) and the A2 emission scenario, and show the projected distribution for the present, 2030, 2050, 2070 and 2100. The CS modeled results show that large areas of southern, western and central regions of Spain are predicted to become climatically more suitable for date palm cultivation and production between 2030 and 2100. The same model indicates that areas of northeast Spain may become suitable for P. dactyliferas’ cultivation between 2050 and 2100. The CS GCM predicts that northern and western Spain will become climatically unsuitable for date palm cultivation by 2100. This model shows roughly 11.5, 14, 19.5 and 28 million hectares will be conducive for date palm growth by 2030, 2050, 2070 and 2100 respectively (Table 1 and Figure 2). The CS model demonstrates that the areas located in 37.5° to 40°N and 1° to 5°W will become highly suitable for date palm cultivation from 2070 to 2100 while in this region the cold stress will remain and impose limitation for date growth until 2070 (Figures 2-6). Using the MR GCM, the model projects large areas in central and southwestern Spain will be climatically suitable for date palm growth by 2100. Additionally, the MR GCM results indicate that some areas in northern Spain may have an opportunity for date palm cultivation. As the climate condition is projected to be suitable by 2100. From the MR GCM, it can be seen that northwestern Spain will be climatically unsuitable, similar to the current situation. This model shows that areas from 37.5° to 40° N and 1° to 5° W and 40° N to 43° N and 4° W to 7° W will become highly conducive for date palm cultivation by 2100 (Figure 3). The MR GCM shows 12.2, 16, 24 and 33.8 million ha of Spain may have suitable climatic conditions for date palm cultivation by 2030, 2050, 2070 and 2100, respectively.
| Years | Projected by CS(GCM) | Projected by MR(GCM) | CS+Slope+land use(Milion ha) | MR + Slope + land use(Milion ha) |
| 2030 | 11.55 | 12.2 | 7.38 | 7.87 |
| 2050 | 13.77 | 15.88 | 8.80 | 10.34 |
| 2070 | 19.49 | 24.12 | 12.81 | 15.14 |
| 2100 | 18.12 | 33.80 | 18.72 | 22.86 |
Table 1: Table 1- Results of CS and MR GCMs and the refined outputs by the location of unsuitable land uses and area less than 10? slopes for date palm cultivation for 2030, 2050, 2070 and 2100.
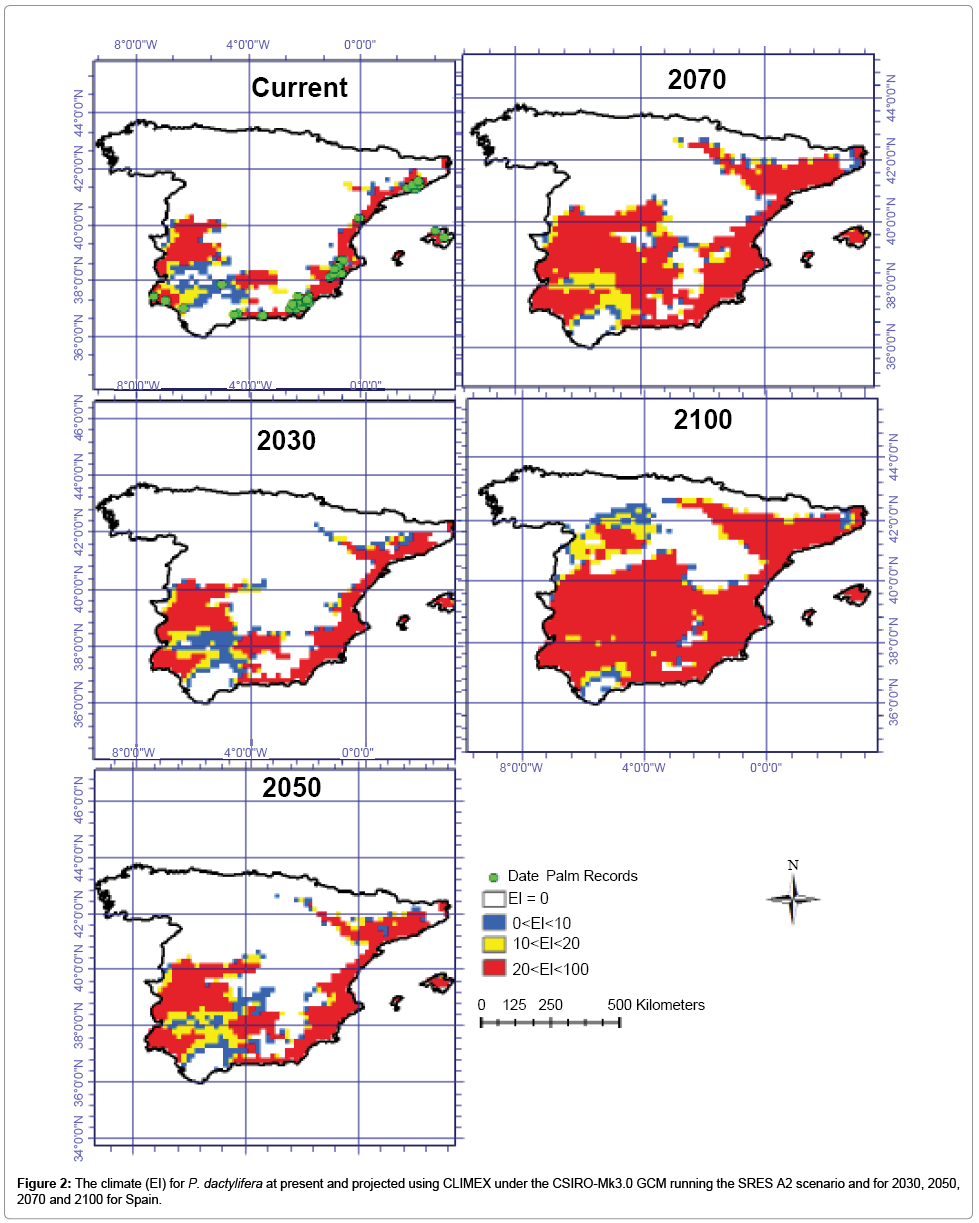
Figure 2: The climate (EI) for P. dactylifera at present and projected using CLIMEX under the CSIRO-Mk3.0 GCM running the SRES A2 scenario and for 2030, 2050, 2070 and 2100 for Spain.
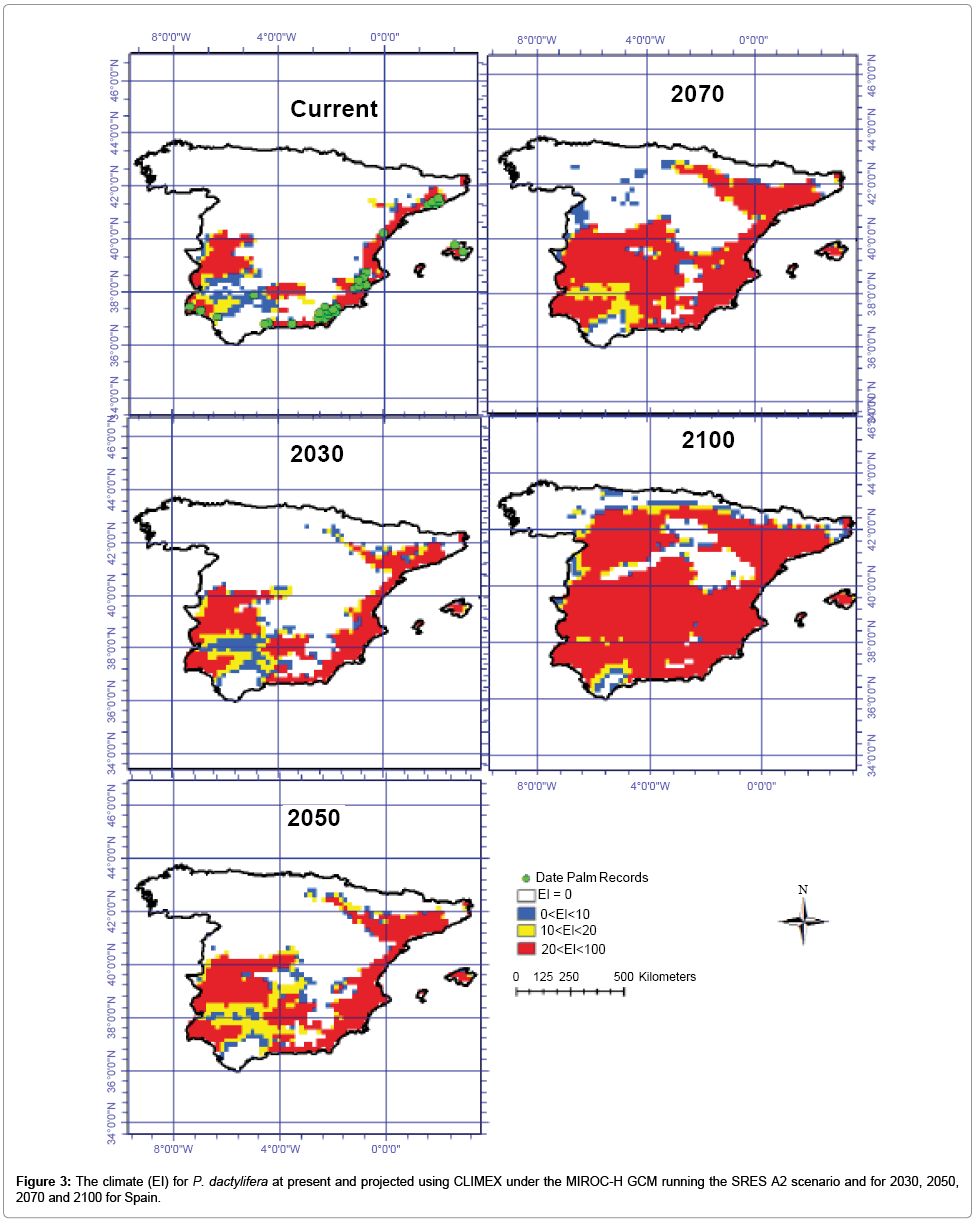
Figure 3: The climate (EI) for P. dactylifera at present and projected using CLIMEX under the MIROC-H GCM running the SRES A2 scenario and for 2030, 2050, 2070 and 2100 for Spain.
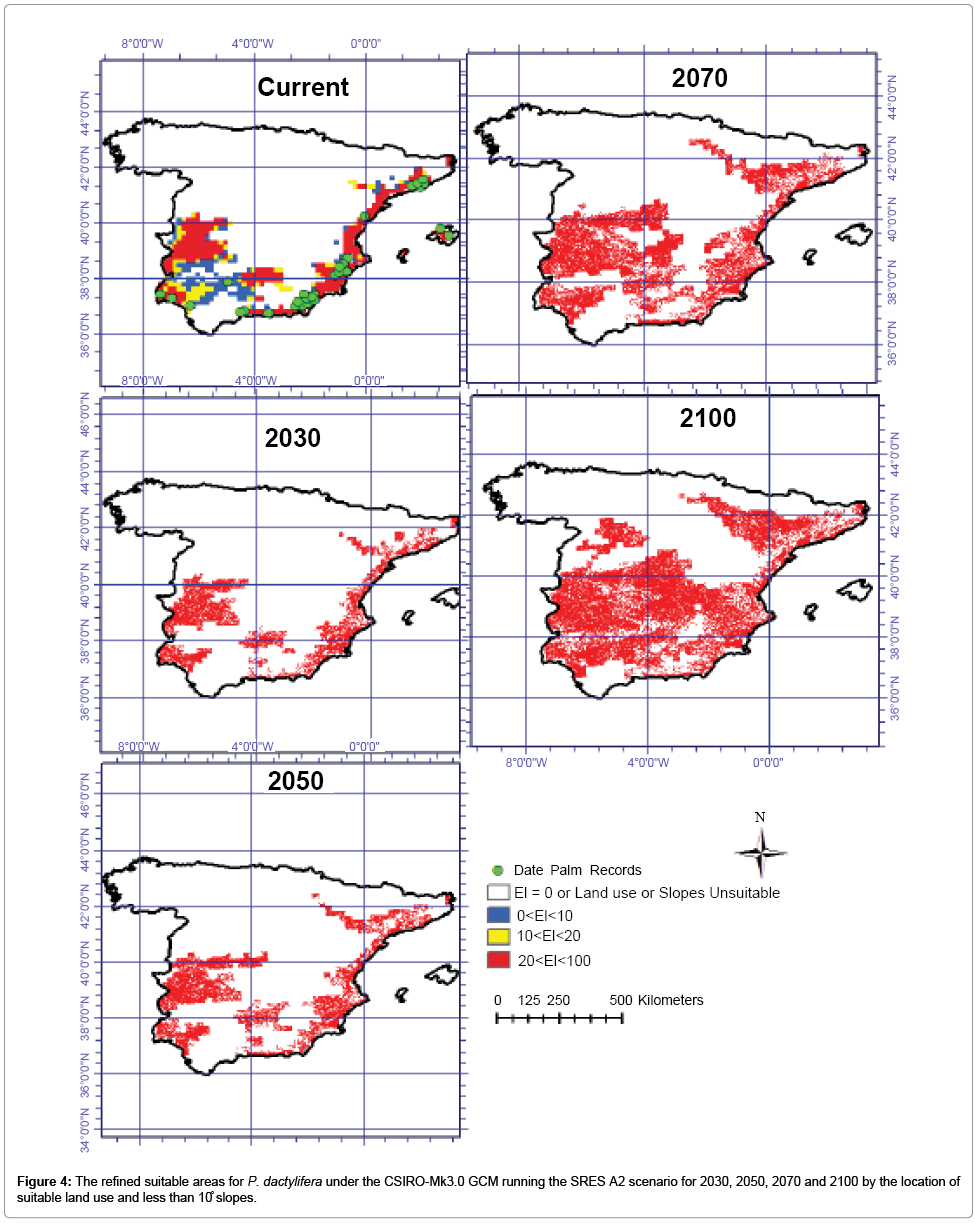
Figure 4: The refined suitable areas for P. dactylifera under the CSIRO-Mk3.0 GCM running the SRES A2 scenario for 2030, 2050, 2070 and 2100 by the location of suitable land use and less than 10̊ slopes.
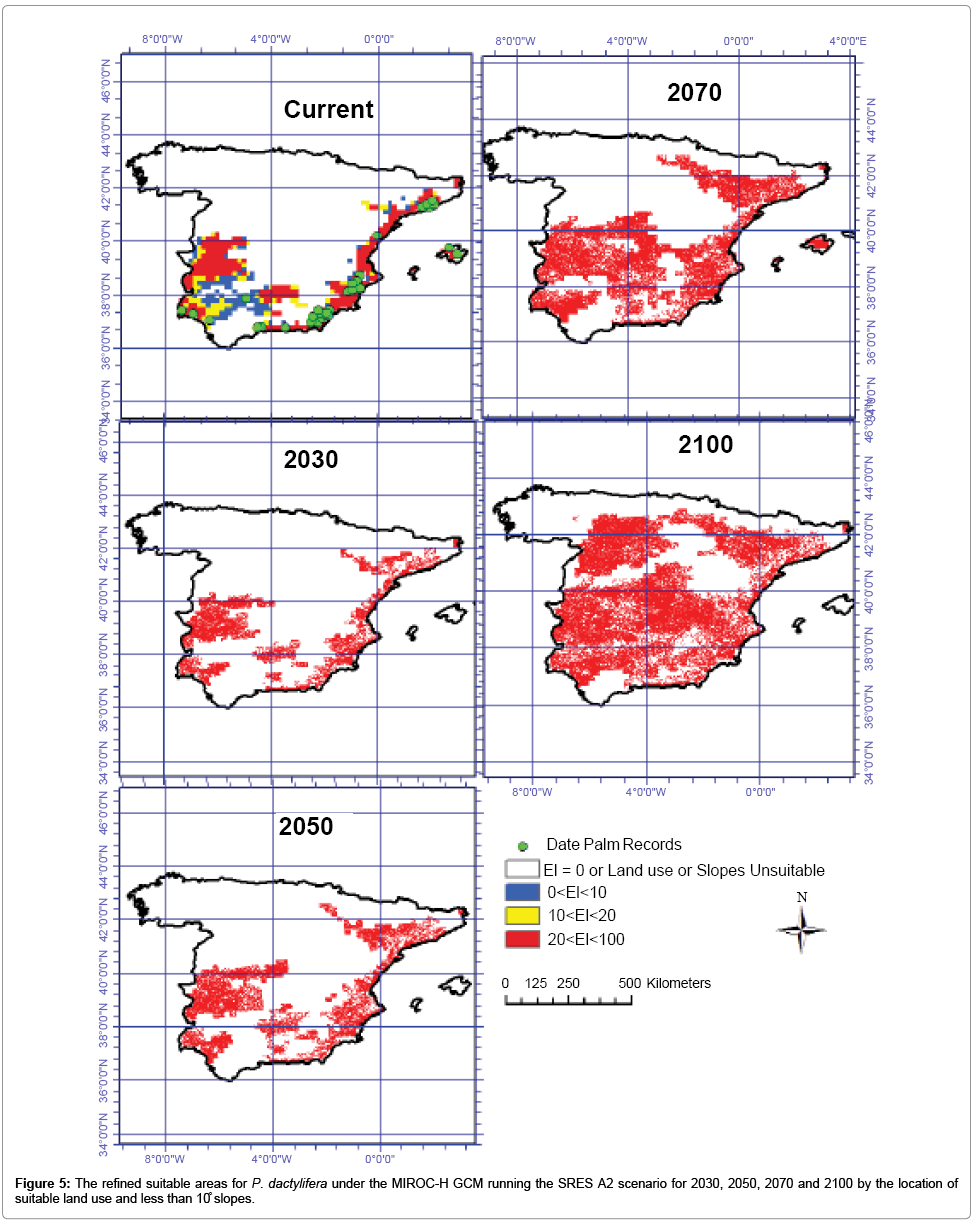
Figure 5: The refined suitable areas for P. dactylifera under the MIROC-H GCM running the SRES A2 scenario for 2030, 2050, 2070 and 2100 by the location of suitable land use and less than 10̊ slopes.
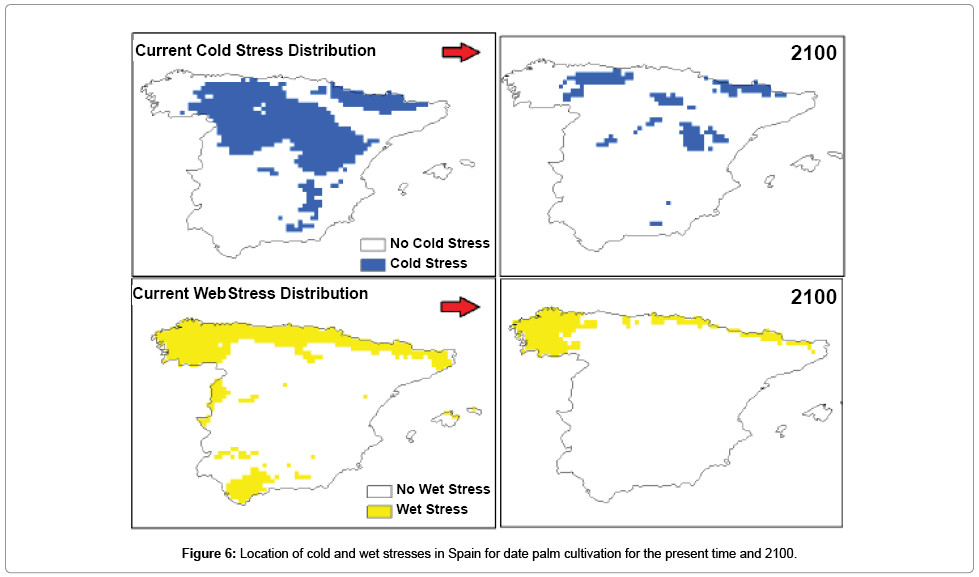
Figure 6: Location of cold and wet stresses in Spain for date palm cultivation for the present time and 2100.
Generally, both CSIRO-Mk3.0 and MIROC-H GCMs project similar trends for Spain. However, some differences can be seen in the projections. These differing results are due to the different predictions of future climate by the two GCMs (Figures 2 and 3 and Table 1) [86,87].
Future climate + Refined results
The refined CS outputs by areas less than 10°slope and the location of suitable land use for date palm cultivation shows that only about 7.38, 8.80, 12.81 and 18.72 million ha will be highly suitable for date plantation by 2030, 2050, 2070 and 2100 in Spain. The refined MR results show that about 7.87, 10.34, 15.14 and 22.86 million ha will be highly conducive for this species growth (Table 1).
Discussion
Species distribution modeling was used as a tool with the aim of illustrating new conducive regions for date palm cultivation in Spain in the future. The results showed that, under current climate, large parts of eastern and western Spain are highly conducive to date palm cultivation (Figure 2), as there are no cold and wet stresses in those regions under the current climate (Figure 6).
There was some agreement in the projections of suitable areas for date palm cultivation between the CS and MR GCMs for Spain. Both MR and CS GCMs project that large parts of Spain will become more climatically suitable for date palm cultivation towards 2100 as a result of significant reduction in areas where currently cold and wet stresses are the major factors restraining date palm distribution (Figures 2, 3 and 6). For example, both models project that the areas with the highest potential for the establishment of date palm by 2100 are central Spain because cold stress may shift northwards (Figure 6). Additionally, both models projected that this unsuitability may be limited to the northern and northwestern regions of Spain due to significant increases in wet stress (Figure 6).
A comparison of the MR and CS models shows some projection differences. While the MR GCM projected that most regions in central Spain could become climatically suitable for date palm growth, the CS GCM projections showed similar results but with less extreme change and effects of change, especially from 2070 to 2100. It should be noted that some differences in the results of CS and MR GCMs (Figures 2 and 3) are due to differences in future greenhouse emission patterns. In this context, the MR model predicts that temperatures will increase by approximately 4.31°C, while the CS model predicts an increase of only 2.11°C by 2100. While the CS model predicts a 14% decrease in future mean annual rainfall, the MR model predicts a mere 1% reduction [2,86,87].
Climate change modeling results indicate the probable changes in the potential distribution of P. dactylifera in the future. As climate changes, certain areas where P. dactylifera currently occurs may become climatically unsuitable, and the economies of those areas will consequently be affected. The results of this study thus have broad implications for future date palm production in Spain for local farmers and the Spanish government, in terms of the predicted increasing possibilities for cultivating date palm. Predictions suggest there will be more areas available that are unaffected by wet or cold stresses, and thus be more conducive for date palm cultivation. In other words, Spain will be able to cultivate these crops to a larger extent in the future compared with the present.
The necessity of refining the CLIMEX results using the slope factor was due to the observation of high correlation between species distribution and abundance as functions of topography which directly effects moisture and fertility gradients [88,89]. Also, most of the studies on distribution of tree species have shown that topography is one of the most important factors affecting yields by causing problems in air permeability, hydraulic conductivity and root developments [90]. Furthermore, many studies have concluded that transportability of the sediment as a consequence of slope creates diverse microsite depending upon the soil type, geology and latitude [89,91,92].
A comparison between the mean CS GCM output and the mean refined results using slope and land use shows that only about 65% of the CS outputs will be suitable for date palm cultivation because of unsuitability in slopes and land uses. The slope map indicates that 34 million ha of Spain has less than 10° slopes and the Spain land use map showed that 2 million ha has highly unsuitable land use for date palm cultivation. For instance, wetland in Spain covers 1.1 million ha and this cannot be used as suitable land for date palm cultivation. From the MR results, refined using the suitable slope and land use, it can be seen that only 65% of the area which was projected to be climatically suitable may be conducive for date palm cultivation while the rest will be unsuitable due to unsuitable slopes and land uses.
A comparison of the results generated by the two models and then refined by slope and land use showed an average difference of about 15%. In the CLIMEX modeled results, the areas that are commonly projected by both GCMs was about 67%. Once the two modeled outputs were refined utilizing land use and slope data, results showed that 83% of areas projected by the two models overlapped for 2030, 2050, 2070 and 2100 (Figure 4 and 5).
It should be noted that there is a need to take into account additional non-climatic factors such as water access to the CLIMEX modeling output. For example, date palm can definitely cope with drought since it has evolved some strategies including high level of water absorption and widespread root system to protect itself in the face of stressors. However, in some areas date palm could only be cultivated (for fruit production) because it is irrigated. Thus, when there is no rainfall or irrigation, fruit production is greatly diminished. Therefore, water availability is an important non-climatic factor and could be a suitable refinement tool to refine the CLIMEX modeling results.
Conclusion
Such modeling allows us an insight into potential impacts of climate change on agricultural production and enables us to take appropriate steps to minimize negative impacts, as well as to take advantage of some scenarios that may arise in the future. Distribution maps such as those used to illustrate this study can assist in the formulation of production methods for cost-effective agricultural crops of the future. Valuable information on current and future changes of areas suitable for date palm cultivation can be deduced from maps such as those shown in Figures 4 and 5 and may encourage plantation owners and managers to consider the future implications of present day management decisions. The methods utilized in this study have equal application for other agricultural crops. Major factors that contributed to the success of this study were: (a) all variables needed for CLIMEX, including temperature, precipitation, sea level pressure and humidity, were readily available; (b) well-documented historical data on date palm distribution was incorporated; and (c) the popular GCMs chosen for this study were on account of (I) their small horizontal grid spacing and (II) their better representation of observed climate at the local scale, compared with GCMs not used in this study. The limitations of this study should be noted. For example, CLIMEX results are based on the response of a species to climate, and non-climatic parameters such as water availability, biotic interactions, soil type and competition between species have not been taken into account. Thus areas shown as being suitable are only based on future climate, suitable slopes and land uses. Large parts of these areas may be unsuitable due to the unsuitability of soil types. Future research could consider these factors in addition to CLIMEX-based modeling.
References
- Jeffrey S, Harold A (1999) Does global change increase the success of biological invaders? Trends in Ecology and Evolution 14: 135-139.
- Hennessy K, Colman R (2007) Global Climate Change Projections. Available.
- Adams M, Rosenzweig C, Peart M, Ritchie T, McCarl A, et al. (1990) Global climate change and US agriculture. Nature 345: 219-224.
- Mas-Coma S, Valero M, Bargues M (2009) Climate change effects on trematodiases, with emphasis on zoonotic fascioliasis and schistosomiasis. Veterinary Parasitology 163: 264-280.
- Bradley BA, Wilcove DS, Oppenheimer M (2010) Climate change increases risk of plant invasion in the Eastern United States. Biological Invasions 12: 1855-1872.
- Fordham DA, Akçakaya HR, Araújo M, Brook BW (2012) Modelling range shifts for invasive vertebrates in response to climate change. Wildlife Conser Chang Clim: 86.
- Luedeling E, Gebauer J, Buerkert A (2009) Climate change effects on winter chill for tree crops with chilling requirements on the Arabian Peninsula. Clim Chang 96: 219-237.
- Pearson RG, Dawson TP (2003) Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Global Ecology and Biogeography 12: 361-371.
- Scherm H, Sutherst RW, Harrington R, Ingram JSI (2000) Global networking for assessment of impacts of global change on plant pests. Environmental Pollution 108: 333-341.
- Araújo MB, Luoto M (2007) The importance of biotic interactions for modelling species distributions under climate change. Global Eco Biogeogra 16: 743-753.
- Kriticos DJ, Sutherst RW, Brown JR, Adkins SW, Maywald GF (2003) Climate change and the potential distribution of an invasive alien plant: Acacia nilotica ssp indica in Australia. J Appl Eco 40: 111-124.
- Visser ME, Both C (2005) Shifts in phenology due to global climate change: The need for a yardstick. Proceedings of the Royal Society B: Biological Sciences 272: 2561-2569.
- Barney JN, DiTomaso JM (2011) Global Climate Niche Estimates for Bioenergy Crops and Invasive Species of Agronomic Origin: Potential Problems and Opportunities. PLoS ONE 6: e17222
- Chakraborty S, Murray GM, Magarey PA, Yonow T, O'Brien RG, et al. (1998) Potential impact of climate change on plant diseases of economic significance to Australia. Austral Plant Path 27: 15-35.
- McDermott M (2009) Climate Change-Induced Drought Causing Crop Failure, Livestock Problems in Indian Himalayas, India.
- Elshibli S, Elshibli E, Korpelainen H (2009) Date Palm (Phoenix dactylifera L.) Plants under Water Stress: Maximisation of Photosynthetic CO2 Supply Function and Ecotypespecific Response. “Biophysical and Socio-economic Frame Conditions for the Sustainable Management of Natural Resources†Tropentag, Hamburg.
- Jones G, Thornton K (2003) The potential impacts of climate change on maize production in Africa and Latin America in 2055. Global Environ Chang 13: 51-59.
- Shabani F, Kumar L, Taylor S (2012) Climate Change Impacts on the Future Distribution of Date Palms: A Modeling Exercise Using CLIMEX. PLoS ONE 7: e48021.
- Ferry M, Gomez S (2002)The Red Palm Weevil in the Mediterranean Area. J Int Palm Soc 46.
- Gómez-Vidal S, Lopez-Llorca LV, Jansson HB, Salinas J (2006) Endophytic colonization of date palm (Phoenix dactylifera L.) leaves by entomopathogenic fungi. Micron 37: 624-632.
- Tengberg M (2011) Beginnings and early history of date palm garden cultivation in the Middle East. Journal of Arid Environments 5: 1-9.
- Ahmed M, Bouna Z, Lemine F, Djeh T, Mokhtar T, et al. (2011) Use of multivariate analysis to assess phenotypic diversity of date palm ( Phoenix dactylifera L.) cultivars. Scientia Horticulturae 127: 367-371.
- Bokhary H (2010) Seed-borne fungi of date-palm, Phoenix dactylifera L. from Saudi Arabia. Saudi Journal of Biological Sciences 17: 327-329.
- Chao C, Krueger R (2007) The Date Palm (Phoenix dactylifera L.): Overview of Biology, Uses, and Cultivation. Journal of Hortscience 42: 1077-1083.
- Veramendi J, Navarro L (1996) Influence of physical conditions of nutrient medium and sucrose on somatic embryogenesis of date palm. Plant Cell, Tissue Organ Cul 45: 159-164.
- Robinson M, Brown B, Williams C (2012) The date palm in southern Nevada. Nevada: The University of Nevada. 23
- Erskine W, Moustafa A, Ahmad E, Lashine A, Badawi T and et al. Date Palm in the GCC countries of the Arabian Peninsula, Egypt: International Center for Agricultural Research in the Dry Areas.
- Anderson P, Lew D, Peterson A (2003) Evaluating predictive models of species’ distributions: criteria for selecting optimal models. Ecological Modelling 162: 211-232.
- Taylor S, Kumar L, Reid N, Kriticos DJ (2012) Climate Change and the Potential Distribution of an Invasive Shrub, Lantana camara.L. PLoS ONE 7: e35565.
- Brooker RW, Travis JMJ, Clark EJ, Dytham C (2007) Modelling species' range shifts in a changing climate: The impacts of biotic interactions, dispersal distance and the rate of climate change. J Theor Bio 245: 59-65.
- Hughes L (2003) Climate change and Australia: Trends, projections and impacts. Aus Eco 28: 423-443.
- Kriticos DJ, Crossman ND, Ota N, Scott JK (2009) Climate change and invasive plants in South Australia. Canberra: CSIRO Climate Adaptation Flagship 97.
- Walther GR, Post E, Convey P, Menzel A, Parmesan C, et al. (2002) Ecological responses to recent climate change. Nature 416: 389-395.
- Kriticos D, Webber B, Leriche A, Ota N, Macadam I, et al. (2011) Global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods in Ecol Evo 3: 53-64.
- Guisan A, Zimmerman NE (2000) Predictive habitat distribution models in ecology. Eco Model 135: 147-186.
- Pearson RG, Dawson TP, Liu C (2004) Modelling species distributions in Britain: a hierarchical integration of climate and land-cover data. Eco 27: 285-298.
- Soberón J (2007) Grinnellian and Eltonian niches and geographic distributions of species. Ecology Letters 10: 1115-1123.
- Sutherst RW, Maywald G (1985) A computerized system for matching climates in ecology. Agri Eco Environ 13: 281-299.
- Sutherst RW, Baker RH, Coakley SM, Harrington R, Kriticos DJ, et al. (2007) Pests under global change – meeting your future landlords? In: Candadell J, Pataki D, Pitelka L, editors. In Terrestrial ecosystems in a changing world. New York: Springer, Berlin Heidelberg. pp. 211-223.
- Follak S, Strauss G (2010) Potential distribution and management of the invasive weed Solanum carolinense in Central Europe. Weed Res 50: 544-552.
- Kriticos DJ (2006) Release Notes for Ozclim Australian Climate Change Scenarios for use in CLIMEX. CSIRO.
- Poutsma J, Loomans AJM, Aukema B, Heijerman T (2008) Predicting the potential geographical distribution of the harlequin ladybird, Harmonia axyridis, using the CLIMEX model. BioControl 53: 103-125.
- Senaratne K, Palmer WA, Sutherst RW (2006) Use of CLIMEX modelling to identify prospective areas for exploration to find new biological control agents for prickly acacia. Aus J Entomol 45: 298-302.
- Phillips SJ, DudÃk M (2008) Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecogra 31: 161-175.
- Sutherst RW, Maywald G, Kriticos DJ (2007) CLIMEX Version 3: User's Guide. In: Ltd HSSP. Melbourne.
- Webber BL, Yates CJ, Le Maitre DC, Scott JK, Kriticos DJ, et al. (2011) Modelling horses for novel climate courses: insights from projecting potential distributions of native and alien Australian acacias with correlative and mechanistic models. Divers and Distrib 17: 978-1000.
- Rogers DJ, Reid RE, Rogers JJ, Addison SJ (2007) Prediction of the naturalisation potential and weediness risk of transgenic cotton in Australia. Agri Ecos & Environ 119: 177-189.
- Saremi H, Kumar L, Sarmadian F, Heidari A, Shabani F (2011) GIS based evaluation of land suitability: a case study for major crops in Zanjan University region. J Food Agri Environ 9: 741-744.
- Beaumont LJ, Hughes L, Poulsen M (2005) Predicting species distributions: use of climatic parameters in BIOCLIM and its impact on predictions of species’ current and future distributions. Ecol Model 186: 251-270.
- Guisan A, Zimmermann NE (2000) Predictive habitat distribution models in ecology. Ecol Model 135: 147-186.
- Fitzpatrick MC, Weltzin JF, Sanders NJ, Dunn RR (2006) The biogeography of prediction error: why does the introduced range of the fire ant over-predict its native range? Global Ecol Biogeography 16: 24-33.
- Jain S, Al-Khayri J, Dennis V, Jameel M (2011) Date Palm Biotechnology. 1st ed: Springer 743.
- Eshraghi P, Zarghami R, Mirabdulbaghi M (2005) Somatic embryogenesis in two Iranian date palm. African Journal of Biotechnology 4: 1309-1312.
- Shayesteh N, Marouf A (2010) Some biological characteristics of the Batrachedra amydraula Meyrick (Lepidoptera: Batrachedridae) on main varieties of dry and semi-dry date palm of Iran.
- Mahmoudi H, Hosseininia G (2008) Enhancing date palm processing, marketing and pest control through organic culture. J Org Sys 3: 30-39.
- Abbas I, Mouhi M, Al-Roubaie J, Hama N, El-Bahadli A (1991) Phomopsis phoenicola and Fusarium equiseti, new pathogens on date palm in Iraq. Mycological Res 95: 509.
- Auda H, Khalaf Z (1979) Studies on sprout inhibition of potatoes and onions and shelf-life extension of dates in Iraq. J Rad Phy Chem 14: 775-781.
- Heakal MS, Al-Awajy MH (1989) Long-term effects of irrigation and date-palm production on Torripsamments, Saudi Arabia. Geoderma 44: 261-273.
- Al-Senaidy M, Abdurrahman M, Mohammad A (2011) Purification and characterization of membrane-bound peroxidase from date palm leaves (Phoenix dactylifera L). Saudi J Bio Sci 18: 293-298.
- Markhand G (2000) Fruit characterization of Pakistani dates. Date Palm Research Institute.
- Hasan S, Baksh K, Ahmad Z, Maqbool A, Ahmed W (2006) Economics of Growing Date Palm in Punjab, Pakistan. Int J Agri and Bio 8: 1-5.
- Elhoumaizi M, Saaidi M, Oihabi A, Cilas C (2001) Phenotypic diversity of date-palm cultivars (Phoenix dactylifera L.) from Morocco. Genetic Res and Crop Evo 49: 483-490.
- Marqués J, Duran-Vila N, Daròs JA (2011) The Mn-binding proteins of the photosystem II oxygen-evolving complex are decreased in date palms affected by brittle leaf disease. Plant Phy and Biochem 49: 388-394.
- Sutherst R, Floyd RB (1999) Impacts of global change on pests, diseases and weeds in Australian temperate forests.
- Harrington R, Woiwod I, Sparks T (1999) Climate change and trophic interactions. Trends in Eco & amp Evol 14: 146-150.
- Kriticos DJ, Reynaud P, Baker RHA, Eyre D (2012) Estimating the global area of potential establishment for the western corn rootworm (Diabrotica virgiferavirgifera) under rain-fed and irrigated agriculture*. EPPO Bulletin 42: 56-64.
- Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecology Letters 8: 993-1009.
- Kriticos D, Potter K, Alexander N, Gibb A, Suckling D (2007) Using a pheromone lure survey to establish the native and potential distribution of an invasive Lepidopteran. J App Eco 44: 853-863.
- Gordon H, Rotstayn L, McGregor J, Dix M, Kowalczyk E, et al. (2002) The CSIRO Mk3 Climate System Model
- IPCC IPoCC (2007) Climate Change 2007: Synthesis Report. Summary for Policymakers.
- Houghton J, Ding Y, Griggs D, Noguer M, van der Linden P, et al. (2001) Climate change 2001: the science basis
- Jain S (2011) Prospects of in vitro conservation of date palm genetic diversity for sustainable production. Emirates J Food and Agri 23: 110-119.
- NASA (2012) ASTER, Advance Spaceborn Thermal Emission and Reflection Radiometer California Institute of Technology.
- Kriticos DJ, Alexander NS, Kolomeitz S (2006) Predicting the potential geographic distribution of weeds in 2080. In: Preston C, Watts JH, Crossman ND, editors. 15th Australian Weeds Conference. Adelaide, Australia: Weed Management Society of South Australia 27-34.
- Crossman ND, Bryan BA, Cooke DA (2011) An invasive plant and climate change threat index for weed risk management: Integrating habitat distribution pattern and dispersal process. Eco Indic 11: 183-198.
- Svenning JC, Skov F (2007) Could the tree diversity pattern in Europe be generated by postglacial dispersal limitation? Ecol Lett 10: 453-460.
- Chiew F, Kirono D, Kent D, Vaze J (2009) Assessment of rainfall simulations from global climate models and implications for climate change impact on runoff studies. 18th World IMACS Australia 3907-3914.
- Suppiah R, Hennessy K (2007) Australian climate change projections derived from simulations performed for the IPCC 4th Assessment Report. 131-152 .
- Suzaki T, Kume A, Ino Y (2005) Effects of slope and canopy trees on light conditions and biomass of dwarf bamboo under a coppice canopy. J Forest Res 10: 151-156
- Basnet K (1992) Effect of topography on the pattern of trees in tabonuco (Dacryodes excelsa) dominated rain forest of Puerto Rico. Biotropica: 31-42.
- Kravchenko AN, Bullock DG, Boast CW (2000) Joint multifractal analysis of crop yield and terrain slope. Agro J 92: 1279-1290.
- Grattan S (2002) Irrigation water salinity and crop production: ANR Publications.
- Tripler E, Shani U, Mualem Y, Ben-Gal A (2011) Long-term growth, water consumption and yield of date palm as a function of salinity. Agri Water Manage 99: 128-134.
Citation: Shabani F, Kumar L, Esmaeili A (2013) Use of CLIMEX, Land use and Topography to Refine Areas Suitable for Date Palm Cultivation in Spain under Climate Change Scenarios. J Earth Sci Clim Change 4: 145. DOI: 10.4172/2157-7617.1000145
Copyright: ©2013 Shabani F, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.